XiaoMi-AI文件搜索系统
World File Search System启动子

利用昆虫启动子进行基因工程控制蚊媒疾病
摘要:蚊子转基因和基因驱动技术为开发有前途的新型媒介传播疾病预防工具提供了基础,这些工具要么抑制野生蚊子种群,要么降低其传播病原体的能力。许多关于具有强大性别、组织和阶段特异性表达谱的基因的调控 DNA 和启动子的研究支持开发可以控制蚊媒疾病的新工具和策略。尽管可用的调控元件列表很重要,但只有有限的一组可以可靠地驱动时空表达。在这里,我们回顾了我们在蚊子中表达有益基因和其他基因的能力方面的进展,并强调了开发新的蚊虫控制和抗病策略所需的信息。

酵母 PHO5 启动子处核小体结构的有效动力学
摘要染色质动力学由重塑酶介导,在基因调控中起着至关重要的作用,正如在典型模型酿酒酵母 PHO5 启动子中建立的那样。然而,有效的核小体动力学,即启动子核小体配置的轨迹,仍然难以捉摸。在这里,我们通过整合已发表的单分子数据推断出这种动力学,这些数据捕获了从受抑制到完全活跃的 PHO5 启动子状态的多核小体配置,以及其他现有的组蛋白周转和新的染色质可及性数据。我们设计并系统地研究了一类新的“受调节的开关滑动”模型,模拟全局和局部核小体(解)组装和滑动。68,145 个模型中只有 7 个与所有数据吻合良好。所有七个模型都涉及滑动和 N-2 核小体的已知核心作用,但通过调节一个组装而不是解体过程来调节启动子状态转换。这与 PHO5 启动子先前观察结果的常见解释一致,但提出了挑战,并表明染色质通过结合竞争而开放。


Cas9 与细菌基因启动子脱靶结合导致沉默和毒性
源自 Cas9 RNA 引导核酸酶的遗传工具为研究和改造细菌提供了必不可少的能力。虽然在 Cas9 应用于哺乳动物细胞的早期就已注意到脱靶效应的重要性,但由于细菌基因组较小,因此很容易避免 Cas9 在细菌基因组中的脱靶切割。尽管如此,一些研究报告了 Cas9 表达有毒的实验设置,即使使用催化失活的 Cas9 变体 (dCas9)。具体而言,dCas9 在与共享特定 PAM(原间隔区相邻基序)近端序列基序的引导 RNA 复合时具有毒性。在这里,我们证明这种毒性是由 Cas9 与必需基因启动子的脱靶结合引起的,脱靶基因的沉默发生在 PAM 近端序列中仅 4 个 nt 的同一性处。在大肠杆菌和其他肠细菌的各种菌株中进行的筛选表明,有毒向导 RNA 的性质会随着脱靶位置序列的进化而改变。这些结果凸显了 Cas9 可能与细菌基因组中数百个脱靶位置结合,从而导致不良影响。在设计和解释细菌中的 CRISPR-Cas 实验时必须考虑这一现象。
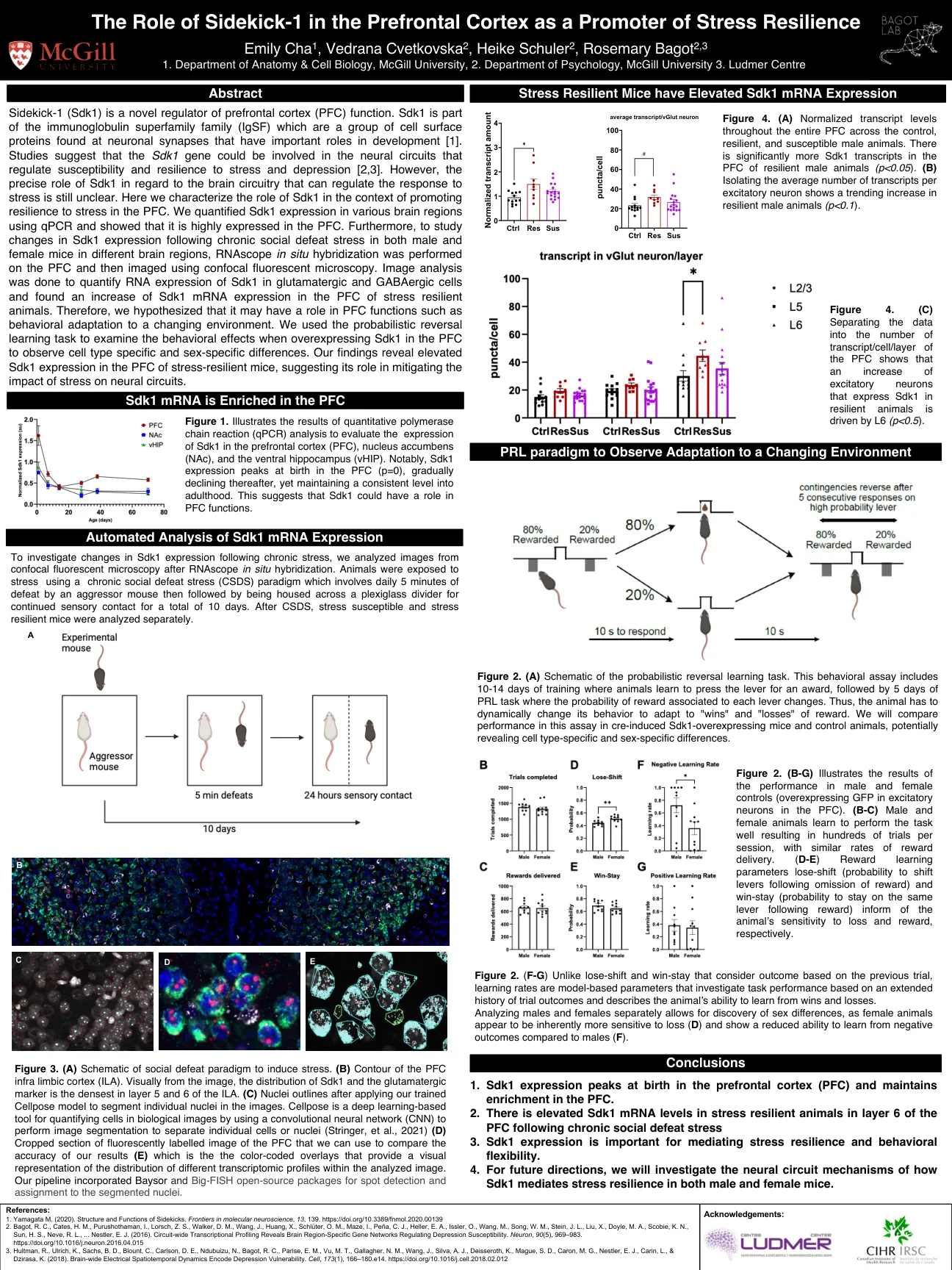
sidekick-1在前额叶皮层中的作用作为应力弹性的启动子
sidekick-1(SDK1)是前额叶皮层(PFC)功能的新型调节剂。SDK1是免疫球蛋白超家族(IGSF)的一部分,它们是在神经元突触中发现的一组细胞表面蛋白,它们在发育中具有重要作用[1]。研究表明,SDK1基因可能参与调节压力和抑郁症的易感性和韧性的神经回路[2,3]。但是,SDK1在可以调节应力反应的脑电路方面的确切作用尚不清楚。在这里,我们表征了SDK1在促进PFC压力的韧性中的作用。我们使用qPCR量化了各个大脑区域中的SDK1表达,并表明它在PFC中高度表达。此外,要研究不同大脑区域的男性和雌性小鼠慢性社交失败压力后SDK1表达的变化,在PFC上进行了原位杂交,然后使用共斑荧光显微镜进行成像。进行图像分析以量化谷氨酸能和GABA能细胞中SDK1的RNA表达,并发现在应激弹性动物的PFC中发现SDK1 mRNA表达增加。因此,我们假设它可能在PFC函数中起作用,例如行为适应不断变化的环境。我们使用概率逆转学习任务来检查PFC中SDK1过表达的行为效应,以观察特定的细胞类型和性别特异性差异。我们的发现显示在应激势力小鼠的PFC中SDK1的表达升高,这表明其在减轻压力对神经回路的影响中的作用。

Marchantia营养发展期间转录因子启动子活动的景观
转录因子(TFS)对于调节基因表达和细胞命运测定至关重要。表征TF基因在时空和时间上的转录活性是了解复杂生物系统的关键步骤。苔藓植物的营养植物分子分生组织具有一些特征,可以与流动植物的芽根尖分生组织具有。然而,与配子植物组织相关的TF的身份和表达方法在很大程度上尚不清楚。只有约450个假定的TF基因,马尔丁塔蒂亚(马丁坦蒂亚多形)是植物系统生物学的出色模型系统。我们已经产生了来自Marchantia TF基因的启动子元素的近乎完整的集合。我们在集合中为所有TF启动子进行了经验测试的记者融合,并系统地分析了Marchantia Gemmae中的表达模式。这使我们能够在早期营养发展中构建表达域的图,并确定一组在干细胞区域中活跃的TF衍生启动子。细胞标记提供了其他工具,并深入了解了配子分生组织的动态调节及其进化。此外,我们为集合中的所有启动子提供了在线表达模式的在线数据库。我们期望这些启动子元素将有助于细胞类型特异性,合成生物学应用和功能基因组学。

靶向具有肿瘤特异性启动子癌细胞基因治疗的策略
癌症是全世界死亡的第二个原因,由于缺乏早期诊断和常规疗法后的高复发率,仅因心血管疾病而超越心血管疾病。化学疗法抑制了癌细胞的快速生长,但它也会以快速增殖速率影响正常细胞。因此,必须制定其他安全,更有效的治疗策略,例如基因治疗,以显着提高癌症患者的生存率和预期寿命。基因疗法的目的是将治疗基因转染到宿主细胞中以表达自身并引起有益的生物学作用。然而,所提出的策略的效率不足以在诊所提供基因治疗的全部潜力。所选择的输送载体类型(病毒或非病毒)取决于基因治疗的所需特定型。第一个基因治疗试验是用由病毒启动子(例如CMV启动子)驱动的治疗基因进行的,CMV启动子除了癌细胞外诱导非特殊毒性的毒性。在肿瘤中使用过表达的肿瘤特异性启动子在给定肿瘤中诱导治疗基因的特定表达,从而增加了其局部活性。已经开发了几个癌症和/或肿瘤特异性启动子系统来靶向癌细胞。基因疗法可用于补充传统疗法以提供更有效的治疗方法。本综述旨在提供有关针对癌症和/或肿瘤特异性启动子靶向基因治疗的最新信息,包括癌症抑制基因,自杀基因,抗肿瘤血管生成,抗肿瘤血管生成,基因沉默和基因编辑技术,以及雇用的运输车辆类型。
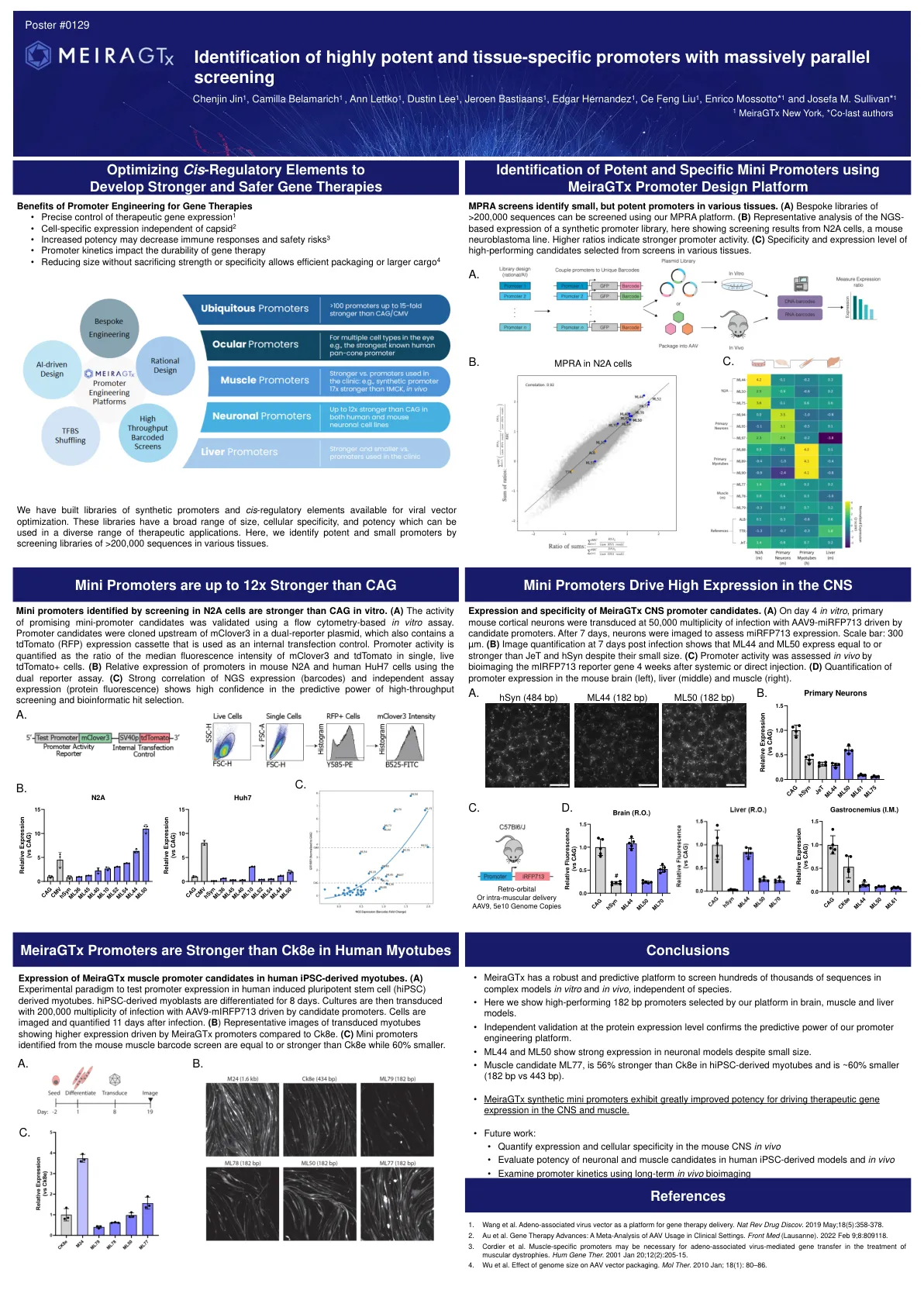
鉴定具有大量平行筛选的高效和组织特异性启动子
迷你启动子在体外比CAG强。(a)使用基于流式细胞术的体外测定法对有希望的迷你启动候选者的活性进行了验证。启动子候选物被克隆在双重孢子质粒中的McLover3上游,该质粒还包含TDTOMATO(RFP)表达盒,该盒被用作内部转染对照。启动子活性被量化为单个活的TDTOMATO+细胞中McLover3和TDTomato的中位荧光强度的比率。(b)使用双报告基因测定法分析,启动子在小鼠N2a和人HuH7细胞中的相对表达。(c)NGS表达(条形码)和独立测定表达(蛋白质荧光)的强相关性表现出对高通量筛选和生物信息学命中选择的预测能力的高信心。

人类基因启动子处的 G-四链体蛋白质相互作用组
基因启动子处的 DNA-蛋白质相互作用在基因表达中起着至关重要的作用。人类细胞的启动子富含富含鸟嘌呤的序列,这些序列可以形成四链 G-四链体 (G4) 结构。G4 正在成为基因调控中一类独特的基于结构的调控元件,它们与蛋白质的相互作用对于 G4 的作用至关重要。目前,我们对 G4-蛋白质相互作用的理解主要是基于个案,没有系统信息。在这项工作中,我们使用来自 ENCODE 项目的数据检查了共识 G4 形成区 G4(+) 周围 1,183 种人类 DNA 结合蛋白(包括转录因子、组蛋白及其修饰酶)的空间占有率。我们发现 G4(+)、其近端侧和远端侧是三个主要的蛋白质结合位点。几乎所有蛋白质在这些位点上都富集或耗尽,这可能是由于竞争或位点之间的时空转换,导致不同程度的变化或持久性,在细胞/组织类型内或跨细胞/组织类型。值得注意的是,组蛋白被排除在 G4(+) 的近端之外,它们与 G4(+) 的结合分别通过乙酰化和甲基化打开和关闭。此外,远端优先富集 H3K23me2 和 H3K4me2。我们的实验还揭示了相应的 G4-蛋白质相互作用模式。总之,我们的结果表明 G4 在动态定义和协调基因启动子处的染色质结构和 DNA-蛋白质相互作用以进行转录调控方面发挥着普遍作用,而这项任务不太可能通过基于序列的 DNA 识别来完成。

分枝杆菌中 CarD 的转录调控受基础启动子动力学指导
细菌病原体,如结核分枝杆菌 ( Mtb ),利用转录因子来使其生理适应宿主内的不同环境。 CarD 是一种保守的细菌转录因子,对 Mtb 的生存至关重要。与通过结合特定 DNA 序列基序来识别启动子的传统转录因子不同, CarD 直接与 RNA 聚合酶结合,以在转录起始期间稳定开放复合中间体 (RP o )。我们之前使用 RNA 测序表明,CarD 能够在体内激活和抑制转录。然而,尽管结合任何 DNA 序列,CarD 如何在 Mtb 中实现启动子特异性调控结果仍不清楚。我们提出了一个模型,其中 CarD 的调控结果取决于启动子的基础 RP o 稳定性,并使用来自具有不同 RP o 稳定性水平的一组启动子的体外转录来测试该模型。我们表明,CarD 直接激活 MTB 核糖体 RNA 启动子 rrnA P3 (AP3) 的全长转录本产生,并且 CarD 的转录激活程度与 RP o 稳定性呈负相关。利用 AP3 的延伸 -10 和鉴别器区域中的靶向突变,我们表明 CarD 直接抑制形成相对稳定 RP o 的启动子的转录。DNA 超螺旋也会影响 RP o 稳定性并影响 CarD 调控的方向,这表明 CarD 活性的结果可受启动子序列以外的因素调控。我们的研究结果为 RNA 聚合酶结合转录因子(如 CarD)如何根据启动子的动力学特性发挥特定的调控结果提供了实验证据。