XiaoMi-AI文件搜索系统
World File Search System启动子

疟疾杂志
摘要背景:明确的启动子是所有生物体遗传研究的基本要素,能够控制内源基因的表达、转基因表达和基因编辑。尽管如此,啮齿动物感染性疟原虫的明确启动子仍然很少。约氏疟原虫尤其如此,它常用于研究疟疾感染的蚊虫阶段和肝脏阶段,以及宿主对感染的免疫反应。方法:从寄生虫的整个生命周期中选择了六个启动子( clag-a 、 dynein heavy chain delta 、 lap4 、 trap 、 uis4 、 lisp2 ),文献中提到这些启动子以阶段特异性的方式控制其基因。还确定了赋予强表达水平的组成型 pybip 启动子的最小启动子长度,这对于报告基因和基因编辑酶的表达很有用。结果:相反,观察到这些启动子赋予了阶段富集基因控制,因为一些寄生虫也有效地在其他阶段使用这些启动子。因此,当单独使用这些启动子时,可能会使启动子交换、阶段靶向重组或基因编辑实验的结果解释复杂化。结论:这些数据表明,实现阶段特异性效应(例如基因编辑)可能最好使用双组分系统,其中独立的启动子活性仅在预期的生命周期阶段重叠。关键词:阶段富集启动子,基因编辑,约氏疟原虫,疟原虫
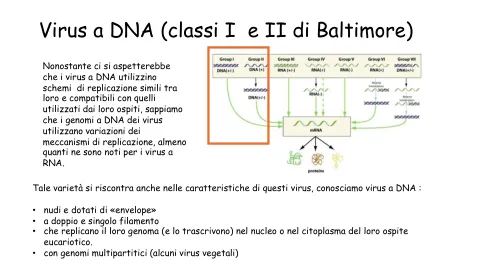
DNA 病毒(巴尔的摩 I 类和 II 类)
• 基因组约为 8 kbp • 它们产生两个同向转录本,其差异剪接产生 8-9 种蛋白质 • 转录可从至少两个启动子开始(P 97、P 670)。第一个是早期基因的启动子,第二个是晚期基因的启动子。 • 来自不同启动子的转录本使用不同的终止子(pAE)• 转录本:• P 97 -pAE 导致蛋白质 E6、7、1、5 的剪接和合成。• P 670 -pALs 导致蛋白质 E4、L1 和 L2 的剪接和合成。 • LCR(长控制区)序列包含与启动子相关的增强子。 • E2 蛋白的完整形式作为启动子(特别是早期启动子)的转录激活剂发挥作用,并与 E1 一起诱导复制(在 LCR 中)。

CRISPR稳定的敲入细胞生产(CAT。C.C408)案例研究:使用CRISPR将红色荧光蛋白(RFP)基因敲入人类胚胎
图2 PCAS-GUIDE-AAVS1和PAAVS1-RFP-DNR的矢量图。pCAS指向AAVS1是哺乳动物细胞中SGRNA和Cas9共表达的多合一载体。SGRNA的表达是由强大的组成型Pol III启动子U6启动子驱动的。而CMV启动子则驱动CAS9酶的表达。paAVS1-RFP-DNR在CMV启动子下的PGK启动子和RFP基因下表达紫霉素的抗性标记。5'和3'AAVS1同源臂(“ aavs-right”和“ aavs-left”)为单元提供了一个用于同源性修复的模板。

设计师基因由人工智能提供
核心启动子不仅确定基因词的启动何方,还决定基础和增强子诱导的条件下的转录活性。在核心启动子内的多个短序列元素已在不同的物种中鉴定出来,但是它们如何共同发挥作用以及它们在何种程度上是真正物种特异性的。在本期的基因和发展中377 - 382)报告合成核心启动子的大规模平行测量值,以生成大量的激活数据集,以告知统计学习模型以识别人和果蝇核心启动子的序列差异。然后将此机器学习模型应用于设计基因核心启动子,这些基因核心启动子特别针对人类转录机械。

新型合成诱导启动子在缺乏基因表达的水缺陷应激中具有转基因杨树中具有绿色组织特异性的基因表达
和3 0区域进行了六聚体,以生成两个较短的合成启动子,Syn3-10b-1(5 0:GTTAACTTCA)和Syn3-10B-2(3 0:GGGCCTGTGG)。将这些启动子的活性与植物中的Syn3进行了比较。syn3和syn3-10b-1在瞬态的农业固定的烟草本nipiana benthamiana叶片中特异性诱导了3天。在稳定的转基因杨树中,Syn3作为本构启动子呈现,但在叶片中的活性最高。SYN3-10B-1在水功率条件下对绿色组织的诱导比模拟对照更强。因此,包含5 0序列的Syn3序列的合成启动子赋予了组织特异性的细胞和水的诱导性转基因杨树,而3 0序列则没有。因此,我们在杨树工程工具包中添加了两个新的合成启动子:Syn3-10B-1,一种绿色组织特异性和水应力诱导的启动子,以及Syn3,Syn3,Syn3,绿色组织预定的构成构成启动子。

ASCL1 和 NEUROD1 依赖基因的启动子是 SCLC 细胞中 lurbinectedin 的特定靶点
小细胞肺癌 (SCLC) 是一种恶性神经内分泌肿瘤,预后较差。本文重点研究神经内分泌 SCLC 亚型 SCLC-A 和 SCLC-N,其转录依赖性由 ASCL 1 和 NEUROD 1 转录因子驱动,这些转录因子靶向 E-box 基序以激活高达 40% 的总基因,根据 ATAC 和 H 3 K 27 Ac 标记,这些基因的启动子保持在稳定开放的染色质环境中。海洋因子 lurbinectedin 利用了这一优势,它优先靶向位于转录起始位点下游的 CpG 岛,从而阻止 RNAPII 延长并促进其降解。这消除了 ASCL 1 和 NEUROD 1 及其依赖基因(如 BCL 2 、 INSM 1 、 MYC 和 AURKA )的表达,这些基因负责相关的 SCLC 致瘤特性(如抑制细胞凋亡和细胞存活)以及其部分神经内分泌特征。总之,我们展示了这些细胞的转录成瘾如何成为它们的致命弱点,以及 lurbinectedin 如何有效地利用这一点作为一种新的 SCLC 治疗手段。

叉质:对异常DNA甲基化对肝细胞癌中替代启动子使用的影响的影响
在发表的文章中,传说中有一个错误的补充图6M,n。使用“启动子活动”而不是“ WGB”进行样品相关聚类。正确的材料语句出现在下面。(M)热图显示了GSE70091中三对启动子活性的相关性。(n)热图显示了删除N3和T3对后,GSE70091中两对启动子活性的性能相关性更好。在已发表的文章中,存在印刷错误。基因名称“ rabgap1l”被错误地写成“ rabgapl1”。对结果进行了校正,甲基化调节的AP可以用作肿瘤诊断标记,第1段。这句话先前指出:“六个MRAP被聚集为四个上调的MRAPS(TNFRSF10的Prmtr.53735,RGS3的Prmtr.32651,CCDC150的Prmtr.36049,RASSF1的Prmtr.5237和RASSF1的Prmtr.5237和Prmtr.5237)和两个下降MRAPS(prmtr.14) prmtr.39585 rabgapl1的启动子活动(图4D,鞋面;表1;表S5)”

医疗政策 - 基因检测 - 恶性神经胶质瘤中MGMT启动子甲基化的分析
在恶性神经胶质瘤中,MGMT(O 6-甲基鸟嘌呤甲基转移酶)基因启动子甲基化的描述/背景测试已被提议作为预测哪些恶性神经胶质瘤患者可能受益于使用烷基化剂化学疗法的方法,例如替莫唑胺(TMZ)。恶性神经胶质瘤通常接受联合治疗,包括切除,化学疗法和放射线。然而,在老年人群中,联合治疗可能太密集了,其中最常见的是这些肿瘤。对这些肿瘤的遗传多样性有了更好的了解,导致努力将分子发现纳入临床实践中,以为包括单药治疗在内的个别患者提供个性化治疗。恶性神经胶质瘤恶性神经胶质瘤是成人最常见的原发性脑癌,在美国,每年约有13,000例新病例。使用世界卫生组织(WHO)组织学标准的脑肿瘤分级对应于恶性(侵略性)的程度,范围从WHO I级(最不侵略性)到IV级(最具侵略性)。 对于恶性神经胶质瘤,间变性星形细胞瘤被认为是III级和胶质母细胞瘤多形(GBM)IV级。 ,GBM是最常见和研究最多的亚型。 1尽管有治疗的进展,但GBM的预后仍然很差,只有三分之一的患者存活了一年,不到5%的患者超过5年。 在2016年,他修订了其中枢神经系统肿瘤(CNS)的分类,以便根据遗传驱动器突变对弥漫性浸润的神经胶质瘤进行分组。使用世界卫生组织(WHO)组织学标准的脑肿瘤分级对应于恶性(侵略性)的程度,范围从WHO I级(最不侵略性)到IV级(最具侵略性)。对于恶性神经胶质瘤,间变性星形细胞瘤被认为是III级和胶质母细胞瘤多形(GBM)IV级。,GBM是最常见和研究最多的亚型。1尽管有治疗的进展,但GBM的预后仍然很差,只有三分之一的患者存活了一年,不到5%的患者超过5年。在2016年,他修订了其中枢神经系统肿瘤(CNS)的分类,以便根据遗传驱动器突变对弥漫性浸润的神经胶质瘤进行分组。2弥漫性神经胶质瘤包括前WHO II和III级星形胶质细胞肿瘤,II级和III少突胶质瘤,IV级胶质母细胞瘤和儿童弥漫性神经胶质瘤。具有胶质母细胞瘤组织学的肿瘤是根据IDH变体的存在分组的。

利用 CRISPR-Cas9 和 Cre-Lox 系统在天然启动子的控制下生成条件突变敲入
通过产生突变来调节基因活性对理解蛋白质功能做出了重大贡献。然而,突变分析通常使用过表达研究,其中蛋白质脱离了其正常的环境和化学计量。在目前的研究中,我们试图开发一种方法,同时使用 CRISPR/Cas9 和 Cre-Lox 技术来修改内源性 SAT1 基因,以引入蛋白质的突变形式,同时仍受其天然基因启动子的控制。我们通过转录终止元件克隆了野生型 (WT) SAT1 的 C 末端部分,并在关键结合位点包含点突变的相同版本 SAT1 前面与 LoxP 位点相邻。在 CRISPR/Cas9 诱导的 DNA 双链断裂后,通过非同源末端连接 (NHEJ) 将构建体插入内源性 SAT1 基因座。在确认插入事件不会改变 SAT1 的正常功能后,我们便能够通过引入 Cre 重组酶来评估点突变的影响。因此,该系统能够生成内源性 WT SAT1 可以有条件地修改的细胞,并允许在正常启动子和染色质调节的背景下研究位点特异性突变的功能后果。