XiaoMi-AI文件搜索系统
World File Search System必需

ESMO网络研讨会:ESMO深水潜水:乳腺癌 - ER+转移性乳腺癌:精炼实践和转向研究2024
•了解分类和风险分层的生物学假设,治疗学领域的持续/必需研究以及使用对乳腺癌的生物标志物精确医学的使用方法的知识。

美国结构化研究股票策略
分析师分配 Tony Wang 信息技术 15.90% Chris Graff 通讯服务 11.05 Ken W. Allen 信息技术 8.96 Jeff Holford 医疗保健 4.90 Gregory Locraft 金融 3.96 Kate Hobbs 医疗保健 3.92 Steve Strycula 必需消费品 3.88 Nina Gupta 金融 3.55 Josepha Kaufman 非必需消费品 3.25 Ari Weisband 金融 3.17 Jill Jortner 医疗保健 2.91 Daniel Shear 通讯服务 2.85 Stephanie Beebe 信息技术 2.26 Vineet S. Khanna 公用事业 2.15 Lee Sandquist 非必需消费品 2.13 Melanie A. Rizzo 工业和商业服务 1.95 Dante Pearson 金融 1.84 Karim Laib 金融 1.73 Jason Adams 工业和商业服务 1.71 Ross MacMillan 信息技术 1.61 Priyar Maniar 能源 1.56 Dhiren Shah 工业和商业服务 1.53 Dinesh Aravindhan 工业和商业服务 1.39 Jason Leblang 工业和商业服务 1.34 Ziad Bakri 医疗保健 1.12 Gregory Korondi 房地产 0.80 Preeta Ragavan 房地产 0.50 Teddy Oaks Financials 0.44

提前离职流程 - MyNavy HR
一旦被接受,水手必须与海军职业顾问合作,通过 MNCC 向 PERS-832 提交 1306/7,并附上 MILPERSMAN 1910-108 中列出的所有必需项目,以申请提前退役。


磁共振引导的集中超声心理切开术 - 临床确定和适应症 - 美国退伍军人事务部
必需震颤是一种神经(神经系统)疾病,会导致身体部位的非自愿摇动或发抖。基本震颤也可能被称为良性基本震颤,是最常见的震颤类型,在任何年龄都可以发生,但在40岁以上的人中最常见。没有已知的原因震颤的原因,但是50%的病例与遗传危险因素有关。研究表明,小脑,丘脑和皮层(大脑结构的组成部分)参与了必需震颤的机制。症状可能包括摇摇欲坠/颤抖的声音,点头点头和握手,影响写作,保持对象或使用工具的能力。症状可能是由某些药物,情绪,疲劳,咖啡因或温度变化引起的。

轻松击败市场的板块轮动策略......
• 材料 (XLB) • 能源 (XLE) • 金融 (XLF) • 工业 (XLI) • 科技 (XLK) • 消费必需品 (XLP) • 公用事业 (XLU) • 医疗保健 (XLV) • 非必需消费品 (XLY)

针对癌症中的谷氨酰胺-精氨酸-脯氨酸代谢轴
摘要 代谢异常是肿瘤的重要特征,谷氨酰胺-精氨酸-脯氨酸轴是肿瘤代谢的重要节点,在氨基酸代谢中起着重要作用,同时也是其他非必需氨基酸和必需代谢物合成的支架。本文就(1)肿瘤细胞对谷氨酰胺的依赖,谷氨酰胺转运和代谢加速;(2)谷氨酰胺进入细胞外、细胞内合成及细胞内谷氨酰胺命运的调控方式;(3)谷氨酰胺、脯氨酸和精氨酸代谢途径及其相互作用;(4)针对谷氨酰胺-精氨酸-脯氨酸代谢系统的肿瘤治疗研究进展作一综述,重点总结了针对该代谢系统关键酶之一P5CS(ALDH18A1)的治疗研究进展,为针对肿瘤代谢特点的治疗提供新的依据。

CMMI 开发,版本 1.3 - SEI 数字图书馆
2 过程域组件 9 核心过程域和 CMMI 模型 9 必需、预期和信息性组件 9 必需组件 9 预期组件 9 信息性组件 10 与第二部分相关的组件 10 过程域 11 目的陈述 11 介绍性说明 11 相关过程域 12 具体目标 12 通用目标 12 具体目标和实践摘要 12 具体实践 13 示例工作产品 13 子实践 13 通用实践 13 通用实践详述 14 附加内容 14 支持性信息性组件 14 注释 14 示例 14 参考文献 15 编号方案 15 印刷约定 16
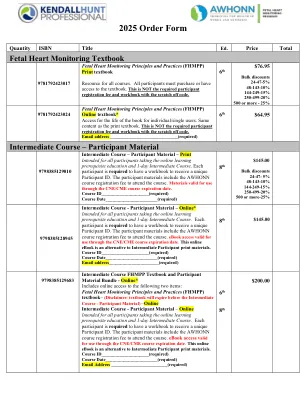
2025订单表格
中级课程FHMPP教科书和参与者材料捆绑包 - 在线访问以下两个项目:胎儿心脏监测原理和实践(FHMPP)教科书 - (免责声明:教科书:中等课程 - 参与者 - 参与者 - 参与者材料) - 参与者 - 在线中间课程 - 参与者 - 在线参与者在线学习的课程和在线学习的预期教育,并进行了前提。每个参与者都必须有一个工作簿才能接收独特的参与者ID。参与者的材料包括参加课程的Awhonn课程注册费。电子书访问v ALID用于通过CNE/CME课程到期日期。本在线电子书是中级参与者印刷材料的替代方法。课程ID _____________________(必需)课程日期_________________________________________________________________________________________________________________________________________________(必需)