XiaoMi-AI文件搜索系统
World File Search System控制分析
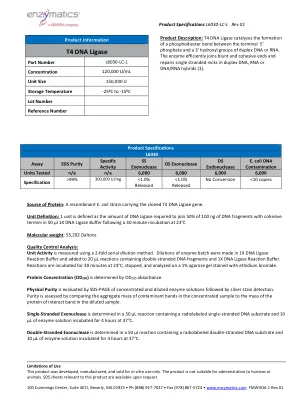
T4 DNA连接酶
蛋白质的来源:带有克隆的T4 DNA连接酶基因的重组大肠杆菌菌株。单位定义:1个单位定义为将100 ng的DNA片段中的50%与粘性末端连接到50 µl 1x 1x DNA连接酶缓冲液后30分钟在23°C分子重量下孵育后所需的50%的DNA片段:55,292 DALTONS质量控制分析:使用2ffliutial serial dilitial doldutial doldiques soge。在1x DNA连接酶反应缓冲液中制作酶批次的稀释液,并添加到含有双束DNA片段和1X DNA连接酶反应缓冲液的20 µL反应中。在23°C下孵育30分钟,停止并在用溴化乙锭染色的1%琼脂糖凝胶上进行分析。蛋白浓度(OD 280)由OD 280吸光度确定。物理纯度,然后进行银色染色检测。通过比较浓缩样品中污染物带的聚集质量与稀释样品中蛋白蛋白蛋白带的质量来评估纯度。单链核酸酶在含有放射性标记的单链DNA底物的50 µL反应中确定,在37°C下孵育4小时4小时。双链外切核酸酶在50 µL反应中确定,该反应含有放射性标记的双链DNA底物和10 µL的酶溶液在37°C下孵育4小时。

通过RP-
摘要目的:为了同时确定散装和药物剂型的达帕格列嗪和Linagliptin,创建并随后验证了一种更容易,更实惠的RP-HPLC方法。方法:使用柱热科学同步C8,5μm粒径,昏暗。(mm)250x4.6 i.D标准化色谱条件。使用波长为230 nm的紫外检测器检测分析物。磷酸盐缓冲液和乙腈在30:70 v/v比的流动阶段使用。结果:Dapagliflozin和Linagliptin的保留时间分别为3.3和2.3分钟。dapagliflozin校准曲线在5–25μg/ml的浓度范围内显示线性相关系数为0.999,而Linagliptin的浓度范围为2-10μg/mL。已经确定,这种方法是快速,健壮,线性,灵敏,准确,精确和特定的。将各种降解应力条件应用于Dapagliflozin和Linagliptin。结论:随着保留时间的明显不同,两种API(活性药物成分)与纯标准药物(Dapagliflozin和Linagliptin)明显区分开。当前的药物方法已经开发并成功地用于确定合并配方和常规质量控制分析中的达帕列酰辛和利格列汀的水平,具有良好的准确性和敏感性。该方法已根据ICH指南对统计验证。关键字:Dapagliflozin,Linagliptin,反相HPLC,验证,强制降解研究,色谱条件。收到:12/09/2024接受:09/10/2024

评估gnotobirotic小鼠和人类中的微生物组指导的零食
*地址为:jgordon@wustl.edu。作者贡献O.D-B。和J.I.G.设计了gnotobiotic小鼠研究。A.C.H. 监督了肥胖人类供体的粪便样品,用于殖民无菌小鼠。 O.D-B。 和N.H.进行了动物研究。 M.J.B.,S.K.,O.D-B。 和J.I.G. 与D.K.H.一起设计了人类研究。和S.V. 谁监督了两种人类研究中使用的纤维零食原型的设计,制造和质量控制分析。 a.m.和S.V. 纤维制剂的有组织的碳水化合物和糖苷连接组成分析。 S.K.监督人类参与者的受控饮食研究。 与K.K.一起 和T.W. J.J.C.,G.C。和C.B.L. 对小鼠饮食和粪便样品进行了质谱测定。 J.C.对从食用2个含有零食的2个和4纤维的参与者那里收集的人类粪便样品进行了LC-QTOF-MS分析。 O.D-B。 监督了小鼠和人类生物测量的存档和处理,并从这些样品中生成了16S rDNA和shot弹枪测序数据集。 M.C.H. 和C.D. 实现了宏基因组装/注释管道。 D.A.R.,S.A.L。和A.O. 进行了粪便微生物的McSeed途径重建,而V.L. 和B.H. 提供了cazyme注释。 A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。A.C.H.监督了肥胖人类供体的粪便样品,用于殖民无菌小鼠。O.D-B。 和N.H.进行了动物研究。 M.J.B.,S.K.,O.D-B。 和J.I.G. 与D.K.H.一起设计了人类研究。和S.V. 谁监督了两种人类研究中使用的纤维零食原型的设计,制造和质量控制分析。 a.m.和S.V. 纤维制剂的有组织的碳水化合物和糖苷连接组成分析。 S.K.监督人类参与者的受控饮食研究。 与K.K.一起 和T.W. J.J.C.,G.C。和C.B.L. 对小鼠饮食和粪便样品进行了质谱测定。 J.C.对从食用2个含有零食的2个和4纤维的参与者那里收集的人类粪便样品进行了LC-QTOF-MS分析。 O.D-B。 监督了小鼠和人类生物测量的存档和处理,并从这些样品中生成了16S rDNA和shot弹枪测序数据集。 M.C.H. 和C.D. 实现了宏基因组装/注释管道。 D.A.R.,S.A.L。和A.O. 进行了粪便微生物的McSeed途径重建,而V.L. 和B.H. 提供了cazyme注释。 A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。O.D-B。和N.H.进行了动物研究。M.J.B.,S.K.,O.D-B。 和J.I.G. 与D.K.H.一起设计了人类研究。和S.V. 谁监督了两种人类研究中使用的纤维零食原型的设计,制造和质量控制分析。 a.m.和S.V. 纤维制剂的有组织的碳水化合物和糖苷连接组成分析。 S.K.监督人类参与者的受控饮食研究。 与K.K.一起 和T.W. J.J.C.,G.C。和C.B.L. 对小鼠饮食和粪便样品进行了质谱测定。 J.C.对从食用2个含有零食的2个和4纤维的参与者那里收集的人类粪便样品进行了LC-QTOF-MS分析。 O.D-B。 监督了小鼠和人类生物测量的存档和处理,并从这些样品中生成了16S rDNA和shot弹枪测序数据集。 M.C.H. 和C.D. 实现了宏基因组装/注释管道。 D.A.R.,S.A.L。和A.O. 进行了粪便微生物的McSeed途径重建,而V.L. 和B.H. 提供了cazyme注释。 A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。M.J.B.,S.K.,O.D-B。和J.I.G.与D.K.H.一起设计了人类研究。和S.V.谁监督了两种人类研究中使用的纤维零食原型的设计,制造和质量控制分析。a.m.和S.V.纤维制剂的有组织的碳水化合物和糖苷连接组成分析。S.K.监督人类参与者的受控饮食研究。 与K.K.一起 和T.W. J.J.C.,G.C。和C.B.L. 对小鼠饮食和粪便样品进行了质谱测定。 J.C.对从食用2个含有零食的2个和4纤维的参与者那里收集的人类粪便样品进行了LC-QTOF-MS分析。 O.D-B。 监督了小鼠和人类生物测量的存档和处理,并从这些样品中生成了16S rDNA和shot弹枪测序数据集。 M.C.H. 和C.D. 实现了宏基因组装/注释管道。 D.A.R.,S.A.L。和A.O. 进行了粪便微生物的McSeed途径重建,而V.L. 和B.H. 提供了cazyme注释。 A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。受控饮食研究。与K.K.一起和T.W.J.J.C.,G.C。和C.B.L. 对小鼠饮食和粪便样品进行了质谱测定。 J.C.对从食用2个含有零食的2个和4纤维的参与者那里收集的人类粪便样品进行了LC-QTOF-MS分析。 O.D-B。 监督了小鼠和人类生物测量的存档和处理,并从这些样品中生成了16S rDNA和shot弹枪测序数据集。 M.C.H. 和C.D. 实现了宏基因组装/注释管道。 D.A.R.,S.A.L。和A.O. 进行了粪便微生物的McSeed途径重建,而V.L. 和B.H. 提供了cazyme注释。 A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。J.J.C.,G.C。和C.B.L.对小鼠饮食和粪便样品进行了质谱测定。J.C.对从食用2个含有零食的2个和4纤维的参与者那里收集的人类粪便样品进行了LC-QTOF-MS分析。O.D-B。 监督了小鼠和人类生物测量的存档和处理,并从这些样品中生成了16S rDNA和shot弹枪测序数据集。 M.C.H. 和C.D. 实现了宏基因组装/注释管道。 D.A.R.,S.A.L。和A.O. 进行了粪便微生物的McSeed途径重建,而V.L. 和B.H. 提供了cazyme注释。 A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。O.D-B。监督了小鼠和人类生物测量的存档和处理,并从这些样品中生成了16S rDNA和shot弹枪测序数据集。M.C.H. 和C.D. 实现了宏基因组装/注释管道。 D.A.R.,S.A.L。和A.O. 进行了粪便微生物的McSeed途径重建,而V.L. 和B.H. 提供了cazyme注释。 A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。M.C.H.和C.D.实现了宏基因组装/注释管道。D.A.R.,S.A.L。和A.O. 进行了粪便微生物的McSeed途径重建,而V.L. 和B.H. 提供了cazyme注释。 A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。D.A.R.,S.A.L。和A.O.进行了粪便微生物的McSeed途径重建,而V.L.和B.H.提供了cazyme注释。A.S.R. 开发了HOSVD和R.Y.C. O.D-B。 和R.A.B. 分析了数据。A.S.R.开发了HOSVD和R.Y.C.O.D-B。 和R.A.B. 分析了数据。O.D-B。和R.A.B.分析了数据。应用于由小鼠和人类生成的数据集的CC-SVD分析平台。对人类研究产生的血浆蛋白质组数据集进行了COMPBIO分析。o.d-b。,C.D.,M.J.B。和J.I.G.O.D-B。 和J.I.G. 在合着者提供的协助下写了这篇论文。O.D-B。和J.I.G.在合着者提供的协助下写了这篇论文。

先进细胞治疗的生产和质量控制指南
农业、畜牧业和食品供应部 - MAPA,法令编号 5,053,2004 年 4 月 22 日。农业、畜牧业和食品供应部 - MAPA,MAPA 监管指令编号 11,2005 年 6 月 8 日。农业、畜牧业和食品供应部 - MAPA,MAPA 监管指令编号 13,2003 年 10 月 3 日。Radrizzani M.、Soncin S.、Lo Cicero V.、Andriolo G.、Bolis S.、Turchetto L. 2016。临床人类间充质基质细胞的质量控制分析:ATMP 释放方法。第 19 章。治疗用品管理局 - TGA,2016 年 8 月。咨询:自体细胞和组织产品的监管以及对生物制品分类的拟议后续变化。治疗用品管理局 - TGA,2015 年 3 月。澳大利亚针对 Remestemcel-L(离体成人人类间充质干细胞)的公共评估报告。原组织。 Torre ML Lucarelli E.、Guidi S.、Ferrari M.、Alessandri G.、De Girolamo L.、Pessina A. 和 Ferrero I. 2014.离体扩增间充质基质细胞临床应用的最低质量要求。 http://online.liebertpub.com/doi/10.1089/scd.2014.0299 姚琳,李哲荣,苏伟伦,李艳萍,林梦莲。间充质干细胞在急性碱烧伤角膜伤口愈合中的作用。 PLoS One,7(2),e30842。 2012 年。

rNase抑制剂
蛋白质的特异性RNase源:带有猪RNase抑制剂基因的重组大肠杆菌菌株。单位定义:1个单位定义为抑制50%cytidine 2',3'-循环单磷酸的水解所需的酶量,由5 ng的RNase A(1)抑制。分子量:74,828 Daltons质量控制分析:使用2倍连续性稀释方法确定单位活动。在1x RNase抑制剂反应缓冲液中制成酶的稀释液,并添加到含有1mm胞苷2',3'-环磷酸1mm的1000 µL反应中,其中包含100mm tris-tris-tris-tris-trisate,1mm Edta,1mm Edta,pH 6.5的1X反应缓冲液中的1μgrNase A。在286nm处的吸光度。蛋白浓度(OD 280)由OD 280吸光度的物理纯度确定,通过浓缩和稀释的酶溶液的SDS-PAGE,然后是银色染色检测。通过比较浓缩样品中污染物带的聚集质量与稀释样品中蛋白蛋白蛋白带的质量来评估纯度。单链核酸酶在含有放射性标记的单链DNA底物的50 µL反应中确定,在37°C下孵育4小时4小时。双链外切核酸酶在50 µL反应中确定,该反应含有放射性标记的双链DNA底物和10 µL的酶溶液在37°C下孵育4小时。双链核酸内切酶在50 µL反应中确定,该反应含有0.5 µg质粒DNA和10 µL的酶溶液在37°C下孵育4小时。
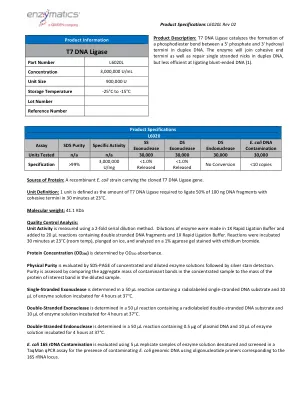
T7 DNA连接酶
蛋白质的来源:一种带有克隆的T7 DNA连接酶基因的重组大肠杆菌菌株。单位定义:1个单位定义为在30分钟内在23°C下在30分钟内将100 ng DNA片段的50%结合的T7 DNA连接酶的量。分子量:41.1 kDa质量控制分析:使用2倍连续稀释方法测量单位活动。稀释液,并添加到含有双链DNA片段和1倍快速连接缓冲液的20 µL反应中。在23°C(室温)下孵育30分钟,浸在冰上,并在用溴化乙锭染色的1%琼脂糖凝胶上进行分析。蛋白浓度(OD 280)由OD 280吸光度确定。物理纯度,然后进行银色染色检测。通过比较浓缩样品中污染物带的聚集质量与稀释样品中蛋白蛋白蛋白带的质量来评估纯度。单链核酸酶在含有放射性标记的单链DNA底物的50 µL反应中确定,在37°C下孵育4小时4小时。双链外切核酸酶在50 µL反应中确定,该反应含有放射性标记的双链DNA底物和10 µL的酶溶液在37°C下孵育4小时。双链核酸内切酶在50 µL反应中确定,该反应含有0.5 µg质粒DNA和10 µL的酶溶液在37°C下孵育4小时。大肠杆菌16S rDNA的污染是使用5 µL r菌酸溶液的样品变性的样品,并在Taqman QPCR分析中筛选,以使用与16S rRNA locus相应的寡核苷酸引物,使用污染的大肠杆菌Genomic DNA。

大脑连接组学和行为研究
摘要背景:自闭症谱系障碍 (ASD) 的异质性阻碍了生物标志物的开发,从而推动了亚型划分工作。大多数亚型划分研究将患有 ASD 的个体分为不重叠的 (分类) 亚组。然而,ASD 的持续个体间变异表明需要采用维度方法。方法:采用贝叶斯模型将患有 ASD 的个体的静息状态功能连接 (RSFC) 分解为多个异常 RSFC 模式,即分类亚型,以下称为“因素”。重要的是,该模型允许每个个体以不同程度表达一个或多个因素 (维度亚型划分)。该模型应用于来自两个多站点存储库的 306 名患有 ASD 的个体 (5.2 – 57 岁)。事后分析将因素与症状和人口统计学联系起来。结果:分析得出三个因素,具有可分离的全脑低 RSFC 和高 RSFC 模式。大多数参与者表达了多个(分类)因素,表明个体内存在多种亚型。所有因素都具有涉及默认模式网络的异常 RSFC,但不同因素之间的方向性(RSFC 过低或过高)不同。因素 1 与核心 ASD 症状有关。因素 1 和 2 与不同的合并症症状有关。年龄较大的男性参与者优先表达因素 3。这些因素在控制分析中表现稳健,与智商或头部运动无关。结论:至少存在三个具有可分离的全脑 RSFC 模式、行为和人口统计学特征的 ASD 因素。因素间异质性默认模式网络 RSFC 过低和过高可能解释了先前报告的不一致。这些因素区分了核心 ASD 和合并症症状——这是 ASD 中一个不太受重视的异质性领域。这些因素在不同程度的 ASD 患者中共同表达,从而协调了 ASD 异质性的分类和维度视角。

系统生物学基础
系统生物学旨在从系统层面理解生物系统。由于多个领域的进步,它是生物学中一个不断发展的领域。最关键的因素是分子生物学的快速进步,以及对 DNA 序列、基因表达谱、蛋白质-蛋白质相互作用等进行全面测量的技术。随着生物数据流的不断增加,现在几乎可以认真尝试将生物系统理解为系统。处理这种高通量实验数据对计算机科学提出了很高的要求,包括数据库处理、建模、模拟和分析。半导体技术的显著进步带来了能够支持系统级分析的高性能计算设施。这不是第一次进行系统级分析的尝试;过去曾有过几项努力,其中最引人注目的是诺伯特·维纳在30多年前提出的控制论或生物控制论。由于当时对分子水平的生物过程的理解有限,大多数工作都是对生理过程的现象学分析。也有生化方法,如代谢控制分析,虽然仅限于稳态流,但它已成功用于探索生物代谢的系统级特性。系统生物学与所有其他新兴科学学科一样,建立在多种共享愿景的努力之上。然而,系统生物学与过去的尝试不同,因为我们第一次能够基于分子水平的理解在系统水平上理解生物学,并创建一个以分子水平为基础的一致知识体系。另外,需要注意的是,系统生物学是系统级研究的生物学,而不是试图将某些教条原则应用于生物学的物理学、系统科学或信息学。当该领域在未来几年成熟时,系统生物学将被描述为系统级生物学领域,广泛使用尖端技术和高度自动化的高通量精密测量,结合复杂的计算工具和分析。系统生物学显然包括实验和计算或分析研究。然而,系统生物学并不是分子生物学和计算科学的简单结合来逆转

lambda exoniclease
X8030测定SDS纯度特异性活性DS核酸内切核蛋白酶DNA污染单位测试了N/A N/A N/A 1500 1500规格> 99%80,000 U/mg NOCONCONION <10份蛋白质来源:从大肠杆菌中纯化的大肠杆菌菌株过表达外生核酸内核酸内核酸酶源于bactreperepteriperophage lambda。单位定义:1个单位定义为在37°C下30分钟内从双链底物中产生10 nmol的酸性脱氧核糖核苷酸所需的酶量。分子量:25.9 kDa质量控制分析:使用2倍连续稀释法测量单位活动。稀释液,并将其添加到含有1.1 kb triTID的DNA片段的50 µL反应中,并加入1x lambda Exo反应缓冲液。在37°C下孵育10分钟,浸入冰上,并使用TCA-PECICTITITART方法进行分析。蛋白浓度(OD 280)由OD 280吸光度确定。物理纯度,然后进行银色染色检测。通过比较浓缩样品中污染物带的聚集质量与稀释样品中蛋白蛋白蛋白带的质量来评估纯度。双链核酸内切酶在50 µL反应中确定,该反应含有0.5 µg质粒DNA和10 µL的酶溶液在37°C下孵育4小时。大肠杆菌16S rDNA的污染是使用5 µL重复的酶溶液的重复样品,并在Taqman QPCR测定中筛选,以使用与16S RRNA locus相应的寡核苷酸引物,以存在污染的大肠杆菌基因组DNA。提供:25毫米Tris-HCl,50 mm NaCl,1 mm DTT,0.1 mm EDTA,50%甘油(25°C时pH 7.5)提供:10x lambda exo反应缓冲液(B8030):670 mm glycine,25 mm mgcl 2(25 mm mgcl 2(pH 9.4)

自然句子阅读过程中情绪处理的神经动力学
阅读时,我们的眼睛通过一系列注视和高速扫视浏览文本,以提取视觉信息。这一过程使大脑能够获得意义,例如关于书面文本中表达的情绪或情感价。大脑在自然阅读过程中如何提取单个单词的情感在很大程度上是未知的。这是由于自然成像的挑战,这导致研究人员之前采用高度控制、定时的逐字呈现缺乏生态效度的定制阅读材料。在这里,我们旨在评估自然阅读英语句子时词语情绪处理的电神经相关性。我们使用了一个公开的数据集,包括同步脑电图 (EEG)、眼动追踪记录和 400 个句子中的 7129 个单词的词级语义注释(苏黎世认知语言处理语料库;Hollenstein 等人,2018 年)。我们计算了注视相关电位 (FRP),即与注视开始时间锁定的诱发电反应。对从视觉和运动诱发活动中清除的 FRP 进行一般线性混合模型分析,结果显示,在注视开始后 224 – 304 毫秒间隔内,左中和右后电极簇中的积极和消极情绪条件之间存在地形差异。包括单词、短语和句子级情绪预测因子的额外分析显示,单词级情绪的 FRP 差异相同,但短语和句子级情绪没有额外的 FRP 差异。此外,从情绪匹配的 40 次试验平均 FRP 中对单词情绪(积极或消极)进行分类的解码分析显示平均准确率为 0.60(95% 置信区间:[0.58, 0.61])。控制分析排除了这些结果是基于眼球运动或语言特征的差异而不是词语情绪。我们的研究结果扩展了以前的研究,表明词汇语义刺激的情感价会在自然阅读过程中对单词注视产生快速的电神经反应。这些结果为在生态有效条件下识别词汇语义处理的神经过程提供了重要的一步,并可用于改进自然语言处理的计算机算法。