XiaoMi-AI文件搜索系统
World File Search System操纵子

通过模拟 CRISPR-Cas 序列来设计高功能基因组编辑器
基因编辑有可能解决农业、生物技术和人类健康领域的基本挑战。源自微生物的基于 CRISPR 的基因编辑器虽然功能强大,但在移植到非原生环境(例如人类细胞)时通常会表现出显著的功能权衡。人工智能 (AI) 支持的设计提供了一种强大的替代方案,有可能绕过进化限制并生成具有最佳属性的编辑器。在这里,使用在大规模生物多样性上训练的大型语言模型 (LLM),我们展示了首次使用 AI 设计的可编程基因编辑器成功精确编辑人类基因组。为了实现这一目标,我们通过系统地挖掘 26 兆碱基的组装基因组和元基因组,整理了超过一百万个 CRISPR 操纵子的数据集。我们通过生成自然界中发现的 CRISPR-Cas 家族中 4.8 倍的蛋白质簇数量并为 Cas9 样效应蛋白定制单向导 RNA 序列来展示我们模型的能力。生成的几个基因编辑器与 SpCas9(典型的基因编辑效应器)相比,表现出相当或更好的活性和特异性,同时在序列上相差 400 个突变。最后,我们展示了一个 AI 生成的基因编辑器,称为 OpenCRISPR-1,它表现出与碱基编辑的兼容性。我们公开发布 OpenCRISPR-1,以促进在研究和商业应用中广泛、合乎道德的使用。

细胞壁脂蛋白CD1687在脱氧氯酸诱导的生物膜艰难梭菌中形成的生物膜中充当DNA结合蛋白
细菌病原体建立复发和持续感染的能力经常与它们形成生物膜的能力有关。梭状芽胞杆菌的差异感染具有较高的复发率和复发率,并且假设生物膜参与其致病性和持久性。生物膜通过C.差异仍然很少了解。已经表明,诸如脱氧胆酸(DCA)或甲硝唑诱导生物膜形成的特定分子,但所涉及的机制仍然难以捉摸。在这项研究中,我们描述了C.差异脂蛋白CD1687在DCA诱导的生物膜形成过程中的作用。我们表明,CD1687的表达是CD1685-CD1689基因簇中的操纵子的一部分,由多个转录启动位点控制,有些是响应DCA诱导的。生物膜形成只需要CD1687,而CD1687的过表达足以诱导生物膜形成。使用RNASEQ分析,我们表明CD1687影响转运蛋白和代谢途径的表达,我们通过下拉测定法(包括转运 - 相关的细胞外蛋白)来识别几个潜在的结合伴侣。然后,我们证明了CD1687在C.差异中暴露于表面,并且该定位是DCA诱导的生物膜形成所必需的。鉴于这种定位以及C.差异形成Edna富生物膜的事实,我们确认CD1687以非特定方式结合DNA。因此,我们假设CD1687是通过通过结合EDNA促进细胞与生物纤维矩阵之间的相互作用,是对DCA的下游响应的组成部分。

使用来自孤立环境细菌的新型回试的噬菌体防御和基因组编辑
反发是细菌免疫系统,可通过杀死受感染的宿主来保护细菌种群免受噬菌体的影响。反击通常包含逆转录酶,一个非编码RNA的模板,该模板被部分转录为RT-DNA和毒性效应子。逆转录酶,非编码RNA和RT-DNA复合物隔离了毒性效应子,直到被噬菌体感染触发为止,此时,毒素被释放出来诱导细胞死亡。由于它们在体内产生单链DNA的能力,还设计了回试以在原核生物和真核生物中生产用于基因组编辑的供体模板。然而,当前的实验表征反元的曲目受到限制,大多数回试来自细菌的临床和实验室菌株。为了更好地了解反逆转录生物学和自然多样性,并扩大了基于反逆转录基因组编辑器的当前工具箱,我们开发了一条管道来分离反替补箱及其细菌宿主与各种环境样品的分离。在这里,我们介绍了这些新颖的反词中的六个,每一个都从不同的宿主细菌中分离出来。我们表征了这些重试的完整操纵子,并测试了它们防御大肠杆菌噬菌体小组的能力。对于其中两个重演,我们通过识别负责触发流产感染的噬菌体基因来进一步揭示其防御机理。最后,我们在大肠杆菌中对这些基因组编辑进行了设计,证明了它们在生物技术应用中的潜在用途。

在海洋和陆地环境中,氨氧化古细菌的新颖秩序水平谱系
氨氧化古细菌(AOA)是地球上最普遍,最丰富的古细菌之一,在海洋,陆地和地热生态系统中广泛分布。与海洋和土壤系统相比,地下环境中AOA种群的基因组多样性,生物地理学和进化过程被大量研究。在这里,我们报告了一种新颖的AOA订单candidatus(CA.)硝基瘤,形成了嗜热ca的姐妹谱系。硝基层。宏基因组和16S rRNA基因读取映射表明,在各种地下水环境中,硝基瘤AOA大量存在及其在一系列地热,陆地和海洋栖息地的广泛分布。陆生氮气肌瘤AOA显示使用甲酸盐作为还原剂来源并使用硝酸盐作为替代电子受体的遗传能力。硝基瘤AOA似乎通过水平基因转移从其他中间人群中获得了关键的代谢基因和操纵子,包括编码尿素酶,亚硝酸盐还原酶和V-type ATPase的基因。获得的功能赋予的其他代谢多功能性可能已促进其辐射到各种地下,海洋和土壤环境中。我们还提供了证据表明,这四个AOA命令中的每一个都跨越了海洋和陆地栖息地,这表明主要AOA谱系比以前提出的更复杂的进化史。一起,这些发现建立了AOA的可靠系统基因组框架,并为该全球丰富的功能公会的生态学和适应提供了新的见解。

使用综合性
多个组件部分的长DNA序列的一锅组装是现代合成生物学构建的迅速产生的关键。的一锅组装方法的方法是由短悬垂链接的多个片段(例如金门)取决于准确和公正的连接。迄今为止的连接设计很大程度上取决于使用经验法则和经验成功的使用,而不是有关连接酶保真度和偏见的详细数据。在这项研究中,我们应用了太平洋生物科学单分子实时测序技术来直接测量单个实验中每个可能的5'基础悬垂配对的连接频率。使用IIS类型限制酶BSAI,已应用此综合数据集来预测金门组装(GGA)的准确性。基于连接数据设计的十个片段组件,其连接数据预计会导致高或低的保真度组件。实验结果不仅证实了总体准确性,还确认了观察到的特定不匹配连接误差及其相对频率。数据进一步用于设计LAC操纵子的12-或24-片段组件,这些组件被证明以高忠诚度和效率组装。因此,连接酶保真度数据允许预测高准确的悬垂对套件的设计比经验法则更大的灵活性,即使在定义的编码区域内,也可以在没有天然DNA序列修改的情况下,在高准确的连接点上安装> 20个片段。

RFS+:一种临床适应性和计算高效的策略,用于增强脑肿瘤分割
摘要:从生态和功能的角度来看,Sanfranciscensis是一种酸味微生物群的重要且主导的细菌种类。尽管该物种在全球酸面团中的不同菌株的普遍存在,但仍需要阐明该物种的遗传多样性背后的驱动因素。在这项研究中,从酸面团样品中分离出14 f。sanfranciscensis菌株,以评估代谢性状的遗传多样性和变异。比较了这14个和31个其他菌株(从NCBI数据库获得)基因组。平均而言,基因组大小和GC含量的值分别为1.31 MBP和34.25%。在45 F. sanfranciscensis菌株中,每个菌株中存在162个核心基因和0至51个独特的基因。核心基因的主要功能与核苷酸,脂质转运和氨基酸以及碳水化合物代谢有关。核心基因的大小占14 F. sanfranciscensis菌株的泛基因组大小的41.18%,即0.70 Mbp为1.70 Mbp。参与碳水化合物利用和抗生素耐药性的14个菌株之间存在遗传变异。此外,还注释了与exodysac-achides生物合成相关的基因,包括epsabd,wxz,wzy。IIA型和IE CRISPR-CAS系统,Pediocin PA-1和Lacticin_3147_A1细菌素操纵子也在F. sanfranciscensis中发现。这些发现可以帮助选择理想的F. sanfranciscensis菌株来开发标准化的启动培养物进行酸面团发酵,并期望为消费者提供更高的质量和营养价值。

嗜热蛋白13:在计算机分析中,提供了与细菌素产生有关的基因的新见解
摘要:细菌素是由细菌和古细菌产生的核糖体合成蛋白质毒素的大家族,具有与生产者菌株紧密相关的物种的抗菌活性。抗菌蛋白质化合物与多种应用有关,包括作为食物和医疗用途的病原体抑制剂。在新鲜和发酵食品保存中常用的几种乳酸细菌(LAB)中,嗜热链球菌以其作为酸奶和奶酪的起始培养而闻名。先前的研究描述了嗜热链球菌SFI13中的细菌素嗜热蛋白13,以及编码其作为操纵子的基因,由两个基因(THMA和THMB)组成。然而,大多数细菌素具有复杂的生产系统,该系统涉及编码具有相对特异性功能的专用蛋白质的几个基因。到目前为止,对嗜热蛋白的合成,调节和表达涉及的基因的关注很少。本研究的目的是在计算机基因挖掘中使用,是研究参与嗜热蛋白13产生的调节系统的存在。的结果显示,专用的推定细菌蛋白基因簇(PBGC),该基因与IIB类菌群基因相似。这个新揭示的PBGC也被发现在嗜热链球菌的各种菌株中,为理解与嗜热蛋白的生产有关的机制提供了一种新的视角和见解。
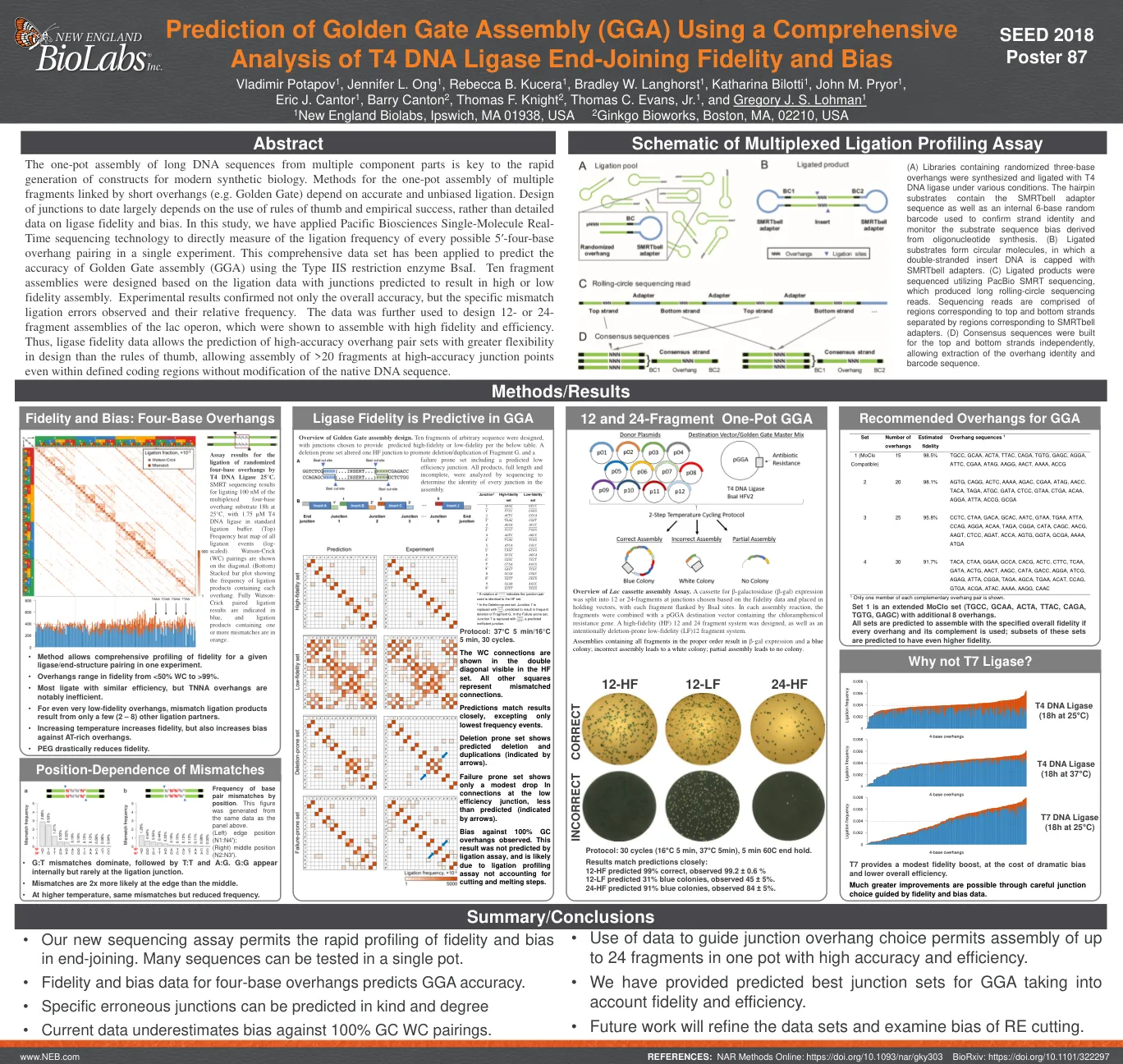
使用综合性
多个组件部分的长DNA序列的一锅组装是现代合成生物学构建的迅速产生的关键。的一锅组装方法的方法是由短悬垂链接的多个片段(例如金门)取决于准确和公正的连接。迄今为止的连接设计很大程度上取决于使用经验法则和经验成功的使用,而不是有关连接酶保真度和偏见的详细数据。在这项研究中,我们应用了太平洋生物科学单分子实时测序技术来直接测量单个实验中每个可能的5'基础悬垂配对的连接频率。使用IIS类型限制酶BSAI,已应用此综合数据集来预测金门组装(GGA)的准确性。基于连接数据设计的十个片段组件,其连接数据预计会导致高或低的保真度组件。实验结果不仅证实了总体准确性,还确认了观察到的特定不匹配连接误差及其相对频率。数据进一步用于设计LAC操纵子的12-或24-片段组件,这些组件被证明以高忠诚度和效率组装。因此,连接酶保真度数据允许预测高准确的悬垂对套件的设计比经验法则更大的灵活性,即使在定义的编码区域内,也可以在没有天然DNA序列修改的情况下,在高准确的连接点上安装> 20个片段。

课程描述植物学
基因组学分类,原核基因组的结构和组织。细菌基因的转录调节剂。细菌基因组中的可转座遗传元素。细菌操纵子和操纵片化的演变。岛屿和致病性和抗性的片段。真核基因组的结构和组织。重复和转座元素及其对基因组的影响。染色体中的端粒和亚电体区域。CpG甲基化和基因沉默。 酵母 - 两种杂交系统。 cDNA微阵列。 线粒体基因组的进化和结构。 基因组测序:整个shot弹枪基因组测序。 测序技术:Sanger毛细血管测序,Roche 454(焦磷酸测序),Illumina/Solexa,固体系统。 测序技术的优缺点。 Maxam-Gilbert测序。 ORF和启动子预测。 内含子和外显子预测。 基因注释。 主要基因组数据库。CpG甲基化和基因沉默。酵母 - 两种杂交系统。cDNA微阵列。线粒体基因组的进化和结构。基因组测序:整个shot弹枪基因组测序。测序技术:Sanger毛细血管测序,Roche 454(焦磷酸测序),Illumina/Solexa,固体系统。测序技术的优缺点。Maxam-Gilbert测序。ORF和启动子预测。 内含子和外显子预测。 基因注释。 主要基因组数据库。ORF和启动子预测。内含子和外显子预测。基因注释。主要基因组数据库。
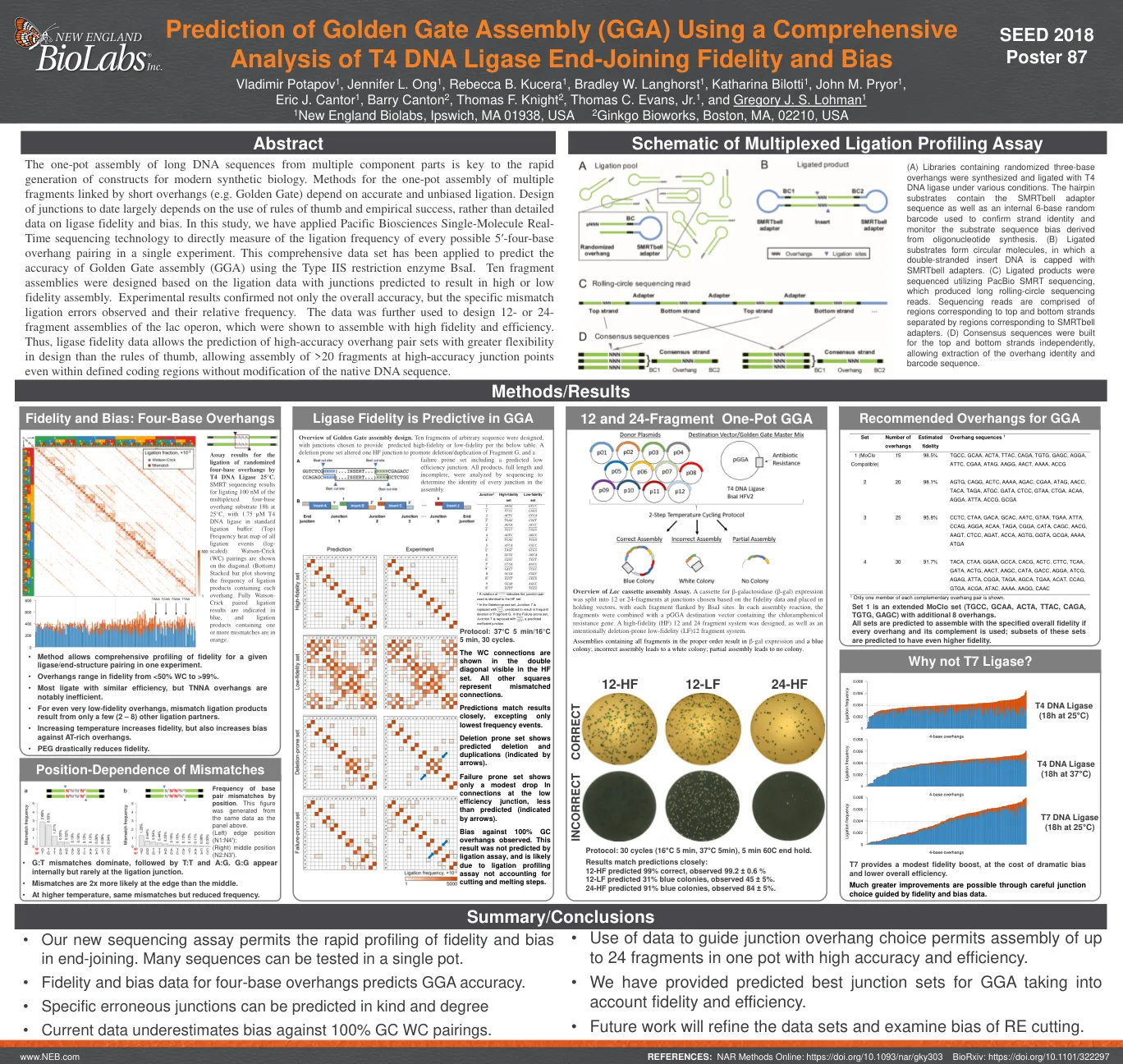
使用综合性
多个组件部分的长DNA序列的一锅组装是现代合成生物学构建的迅速产生的关键。的一锅组装方法的方法是由短悬垂链接的多个片段(例如金门)取决于准确和公正的连接。迄今为止的连接设计很大程度上取决于使用经验法则和经验成功的使用,而不是有关连接酶保真度和偏见的详细数据。在这项研究中,我们应用了太平洋生物科学单分子实时测序技术来直接测量单个实验中每个可能的5'基础悬垂配对的连接频率。使用IIS类型限制酶BSAI,已应用此综合数据集来预测金门组装(GGA)的准确性。基于连接数据设计的十个片段组件,其连接数据预计会导致高或低的保真度组件。实验结果不仅证实了总体准确性,还确认了观察到的特定不匹配连接误差及其相对频率。数据进一步用于设计LAC操纵子的12-或24-片段组件,这些组件被证明以高忠诚度和效率组装。因此,连接酶保真度数据允许预测高准确的悬垂对套件的设计比经验法则更大的灵活性,即使在定义的编码区域内,也可以在没有天然DNA序列修改的情况下,在高准确的连接点上安装> 20个片段。