XiaoMi-AI文件搜索系统
World File Search System极性

关联极性/非极性 LaCrO3/SrTiO3 (001) 界面处的极性畸变和界面电荷
这是作者的同行评审并被接受的稿件。但是,一旦经过编辑和排版,记录的在线版本将与此版本不同。请以 DOI 引用本文:

极性微藻
博士Daniela Morales-Sánchez是TuxtlaGutiérrezInstitute(2005)和Maestra(2007)和Maestra(2007)和Doctor(2014)的生物化学工程师,并在UNAM生物学研究所的生化科学中。 div>他们的博士研究集中于生物柴油生产的异养微藻的培养。 div>在他在美国内布拉斯加州大学的第一个博士后(2014-2017)期间,他的研究导致了通过遗传和代谢工程策略的Chlamydomonas Reinhardtii的脂质含量的增加。 div>2017年,他在挪威的北部大学开始了第二个博士后,在那里他研究了精神噬菌体微藻对产生高价值代谢产物的潜力。 div>目前是ING部的定期调查员。细胞和生物催化,该研究指导与高强度光线适应/适应极性微藻有关的项目,以改善热带微藻。 div>
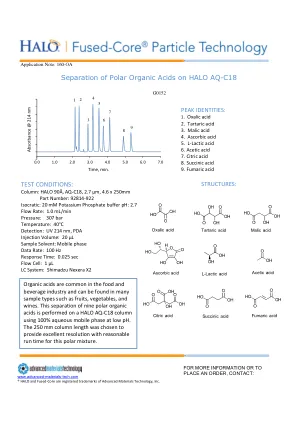
AQ-C18 色谱柱上的极性有机酸
有机酸在食品和饮料行业中很常见,可以在水果、蔬菜和葡萄酒等多种样品中找到。在 HALO AQ-C18 色谱柱上使用低 pH 值的 100% 水相流动相对九种极性有机酸进行分离。选择 250 毫米长的色谱柱可为这种极性混合物提供出色的分辨率和合理的运行时间。


16 bit 模数转换器TM7706 - NET
注释: 1.B 级温度范围为 -40 ℃ ~+85 ℃。 2.这些数据是按最初设计的产品发布的。 3.一次校准实际上是一次转换,因此这些误差就是表 1 和表 3 所示转换噪声的阶数。这 适用于在期望的温度下校准后。 4.任何温度条件下的重新校准将会除去这些漂移误差。 5.正满标度误差包括零标度误差 ( Zero-Scale Error )(单极性偏移误差或双极性零误 差),且既适用于单极性输入范围又适用于双极性输入范围。 6.满标度漂移包括零标度漂移 (单极性偏移漂移或双极性零漂移)且适用于单极性及 双极性输入范围。 7.增益误差不包括零标度误差,它被计算为满标度误差——对单极性范围为单极性偏移 误差,而对双极性范围为满标度误差——双极性零误差。 8.增益误差漂移不包括单极性偏移漂移和单极性零漂移。当只完成了零标度校准时,增 益误差实际上是器件的漂移量。 9.共模电压范围:模拟输入电压不超过 V DD +30mV ,不低于 GND-30mV 。电压低于 GND-200mV 时,器件功能有效,但在高温时漏电流将增加。 10.这里给出的 AIN ( + )端的模拟输入电压范围,对 TM7706 而言是指 COMMON 输入 端。输入模拟电压不应超过 V DD +30mV, 不应低于 GND-30mV 。 GND-200mV 的输入 电压也可采用,但高温时漏电流将增加。 11.VREF=REF IN ( + )- REF IN ( - )。 12.只有当加载一个 CMOS 负载时,这些逻辑输出电平才适用于 MCLK OUT 。 13.+25 ℃时测试样品,以保证一致性。 14.校准后,如果模拟输入超过正满标度 , 转换器将输出全 1, 如果模拟输入低于负满标度, 将输出全 0 。 15.在模拟输入端所加校准电压的极限不应超过 V DD +30mV 或负于 GND - 30mV 。 16.当用晶体或陶瓷谐振器作为器件的时钟源时 (通过 MCLK 引脚 ), V DD 电流和功耗 随晶体和谐振器的类型而变化 (见“时钟和振荡器电路”部分)。 17.在等待模式下,外部的主时钟继续运行, 5V 电压时等待电流增加到 150 μ A , 3V 电 压时增加到 75 μ A 。当用晶体或陶瓷谐振器作为器件的时钟源时,内部振荡器在等待 模式下继续运行,电源电流功耗随晶体和谐振器的类型而变化 (参看“等待模式” 一节)。 18.在直流状态测量,适用于选定的通频带。 50Hz 时, PSRR 超过 120dB (滤波器陷波 为 25Hz 或 50Hz )。 60Hz 时, PSRR 超过 120dB (滤波器陷波为 20Hz 或 60Hz )。 19.PSRR 由增益和 V DD 决定,如下:


极性区域气候模型RACMO2.4
摘要。在本研究中预先提出了极地区域大气气候模型(称为RACMO2.4P1)的下一个版本。主更新包括嵌入Intecast的预测系统(IFS)周期47R1的物理参数包装包。这构成了降水,对流,湍流,气溶胶和表面方案的变化,并包括一种新的云方案,具有更多的预后变量和专用的湖泊模型。fur-hoverore,独立的IF辐射物理模块ECRAD被纳入RACMO,并引入了非冰期区域的多层雪模量。其他更新涉及引入分数陆地面膜,新的和更新的气候数据集(例如气溶胶构成和叶子面积指数),以及对冰川区域的几个参数化的修订。作为概念证明,我们向格陵兰,南极和北极地区的地区展示了第一个结果。通过将结果与观测结果和先前模型版本(RACMO2.33)的输出进行比较,我们表明该模型在表面质量平衡,表面体能平衡,温度,风速,风速,云含量和积雪深度方面很好地形成了。雪水头的对流强烈影响冰盖的局部表面质量平衡,特别是在高积累的地区,例如东南绿地和南极半岛。我们严格评估模型输出,并确定一些可以从进一步的模型开发中拟合的过程。

材料分类的极性金属分类法和...
在过去的十年中,将破裂的反转对称性与金属电导率结合在一起的材料已从思想实验转变为增长最快的研究主题之一。在2013年,在金属3中观察到第一个无可争议的极性转变lioso 3启发了对该受试者的理论和实验性工作的激增,发现了许多材料,这些材料结合了以前被认为是禁忌的特性[nat。mater。12,1024(2013)]。 通常在新生的领域中,兴趣的突然上升伴随着多样化(有时是爆发)术语。 尽管“类似铁电的”金属在理论上是正确确定的,即,在表现出金属电子传输的同时,经历对称性的过渡到极相的材料,但实际材料却发现了多种方法来推动这种定义的边界。 在这里,我们从理论,模拟和实验的角度审查并探索了新兴的极地金属边界,同时引入了统一的分类学。 该框架允许人们描述,识别和分类极性金属;我们还使用它来讨论“铁电”和“金属”一词固有的理论与现实模型之间的一些基本张力。此外,我们强调了静电掺杂模拟在建模极性金属的不同亚类中的缺点,并指出了这种方法的假设如何与实验不同。 我们包括一项已知材料的调查,该调查将极性对称性与金属电导率结合在一起,并根据用于协调这两个顺序及其所得属性的机制进行分类。12,1024(2013)]。通常在新生的领域中,兴趣的突然上升伴随着多样化(有时是爆发)术语。尽管“类似铁电的”金属在理论上是正确确定的,即,在表现出金属电子传输的同时,经历对称性的过渡到极相的材料,但实际材料却发现了多种方法来推动这种定义的边界。在这里,我们从理论,模拟和实验的角度审查并探索了新兴的极地金属边界,同时引入了统一的分类学。该框架允许人们描述,识别和分类极性金属;我们还使用它来讨论“铁电”和“金属”一词固有的理论与现实模型之间的一些基本张力。此外,我们强调了静电掺杂模拟在建模极性金属的不同亚类中的缺点,并指出了这种方法的假设如何与实验不同。我们包括一项已知材料的调查,该调查将极性对称性与金属电导率结合在一起,并根据用于协调这两个顺序及其所得属性的机制进行分类。我们通过使用我们的分类法来描述发现新型极地金属的机会来得出结论。

大脑发育中的细胞内交通和极性
构成人类大脑的神经元是在胚胎发育过程中由神经干细胞和祖细胞通过神经发生过程产生的。导致神经干细胞形态和功能异质性的关键特征是细胞极性,细胞极性被定义为细胞成分的不对称分布。细胞极性的建立和维持得益于极性蛋白和极性产生细胞器(如内质网 (ER) 和高尔基体 (GA))之间的相互作用。ER 和 GA 影响膜成分的分布,并充当将聚糖添加到新生蛋白质和脂质中的枢纽。在过去的几十年里,我们对极性在神经干细胞和祖细胞中的作用的了解大大增加。然而,交通和相关糖基化在神经干细胞和祖细胞中的作用仍然相对未被充分探索。在这篇综述中,我们讨论了细胞极性、结构、身份和细胞内交通之间的联系,并强调了对神经元的研究如何塑造了我们对交通和极性的认识和概念框架。最后,我们将讨论一组称为先天性糖基化障碍 (CDG) 的罕见疾病如何提供独特的机会来研究神经发育背景下的交通和糖基化的贡献。

91 gan中的晶体极性区分...
总而言之,提出的DFT研究表明,在晶状体底物上的N止极gan结构在能量上比GA极极可取。在群集中Ga和N原子的不同可能构型中,仅N止痛器一个是稳定的,而最初的GA极性结构则证明了AB-Initio优化期间的极性变化。DFT建模结果与在硅底物顶部在石墨烯层上生长的GAN纳米线的独家N极性的实验观察一致[2,3]。