XiaoMi-AI文件搜索系统
World File Search System正交
1个国家主要实验室,微生物学院,中国科学院,北京100101,中国 *通信:liwei_zhou1982@im。2江南大学的未来食品科学中心,中国214122 *通信:yanfengliu@jiangnan.edu.cn收到:2024年1月30日; AC
图1。在大肠杆菌的正交线性质粒(O-Replicon)中的正交复制系统的结构和表征不干扰宿主基因组的复制,在大肠杆菌中已成功发展。末端蛋白(TP),正交DNAP(O-DNAP),双链DNA结合蛋白(DSB)和PRD1噬菌体的单链DNA结合蛋白(SSB)的基因在iPTG-诱导促销基因组合的控制下被编码并表达。这些蛋白质与含有倒末端重复序列的异源基因的DNA序列(ITR)相互作用,最终在体内获得正交复制线性质粒。此外,戈伊斯可以通过设计的 - 毒素O-DNAP的连续作用来实现快速而独立的进化,例如在短时间内的短期内显着提高细胞对抗生素的耐药性以及GFP的荧光强度。

连续变化系统中的位移和旋转的量子增强感应
3传感来自正交测量值10 3.1位移感测,并通过正交测量值传感。。。。。。。。。。。。10 3.1.1 fock状态。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。11 3.1.2相干状态。。。。。。。。。。。。。。。。。。。。。。。。。。。。。13 3.1.3高斯州。。。。。。。。。。。。。。。。。。。。。。。。。。。。。14 3.1.4猫状态。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。16 3.1.5 Fock状态叠加。。。。。。。。。。。。。。。。。。。。。。。。18 3.1.6结果摘要。。。。。。。。。。。。。。。。。。。。。。。。。19 3.2旋转传感,并进行正交测量。。。。。。。。。。。。。。。20 3.2.1 fock状态。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。21 3.2.2相干状态。。。。。。。。。。。。。。。。。。。。。。。。。。。。。22 3.2.3高斯州。。。。。。。。。。。。。。。。。。。。。。。。。。。。。23 3.2.4猫状态。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。24 3.2.5结果摘要。。。。。。。。。。。。。。。。。。。。。。。。。24

课程R18
真实对称矩阵L的对角化:6小时正交矩阵 - 对角线形式向对角矩阵的正交转换 - 通过正交转换将二次形式的二次形式还原为规范形式。一阶普通微分方程L:11小时莱布尼兹方程 - 伯努利方程 - 一阶和较高程度的方程 - clairauts形式 - 应用:正交轨迹。高阶线性微分方程L:恒定系数的第二和更高顺序的11小时线性方程 - Euler's and Legendre的线性方程 - 参数变化方法 - 一阶同时线性方程,具有恒定系数 - 应用 - 应用。几个变量的函数L:11小时总导数 - 泰勒的串联扩展 - 两个变量的功能的最大值和最小值 - 受约束的最大值和最小值:Lagrange的乘数方法具有单个约束 - 雅各布人。

书籍(PDF) - 带有Python的高级定量经济学
1个正交预测及其应用5 1.1概述。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。5 1.2关键定义。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。6 1.3正交投影定理。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。9 1.4正式基础。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。1。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。14 1.6回归方。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。15 1.7正交化和分解。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。17 1.8练习。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。。18


放射性核素刺激动态疗法正交靶向破骨细胞和骨髓瘤细胞诱导多维细胞死亡途径
原理:多发性骨髓瘤 (MM) 是一种骨髓浆细胞多灶性恶性肿瘤,其特征是缓解和复发的恶性循环,最终导致死亡。由于骨微环境 (BME) 和 MM 细胞 (MMC) 之间复杂的相互作用,该疾病大多无法治愈。在骨病的“恶性循环”中,MMC 对破骨细胞 (OC) 的异常激活会导致严重的骨溶解、促进免疫逃避并刺激 MMC 的生长。破坏这些癌症-基质相互作用将增强治疗反应。方法:为了打破这种循环,我们将载有非治疗剂量光敏剂二茂钛 (TC) 的纳米胶束 (NM) 正交靶向表达 VLA-4 (α 4β1、CD49d/CD29) 的 MMC (MM1.S) 和表达 α vβ3 (CD51/CD61) 的 OC。同时,全身施用非致死剂量的放射性药物 18 F-氟脱氧葡萄糖 ([ 18 F]FDG) 与 TC (放射性核素刺激疗法,RaST) 相互作用产生细胞毒性活性氧 (ROS)。在 MM1.S 细胞系以及异种移植和同种移植 MM 动物模型中表征了 RaST 的体外和体内作用。结果:我们的数据显示,RaST 诱导细胞脂质的非酶氢过氧化,最终导致线粒体功能障碍、DNA 碎片化和 MMC 的 caspase 依赖性凋亡,使用 VLA-4 亲和 TC-NMs。RaST 上调了 BAX、Bcl-2 和 p53 的表达,突出了通过 BAK 非依赖性途径诱导细胞凋亡。多铜氧化酶 F5 表达的增强(可抑制脂质氢过氧化和 Fenton 反应)不足以克服 RaST 诱导的不可逆功能扰乱 α,β-醛积累增加,这些醛会对 DNA 和蛋白质造成严重且持久的损害。在体内,VLA-4-TC-NM 或 α vβ3-TC-NMs RaST 均对免疫功能低下但免疫功能不正常的 MM 携带小鼠模型产生严重治疗效果。VLA-4-TC-NM 和 α vβ3-TC-NMs 联合治疗可协同抑制骨溶解、减轻肿瘤负担并防止两种 MM 体内模型中的快速复发。结论:通过同时靶向 MM 和骨细胞,联合 RaST 通过对骨癌恶性循环的多管齐下的作用抑制 MM 疾病进展。我们的工作没有采用标准的多药疗法,而是揭示了一种独特的光物理治疗模式,即使用无毒剂量的单一光敏药物正交地作用于癌症和骨细胞,然后通过放射性核素刺激产生 ROS 来抑制肿瘤进展并最大限度地减少免疫功能正常的小鼠和免疫功能低下的人类 MM 模型中的骨溶解。
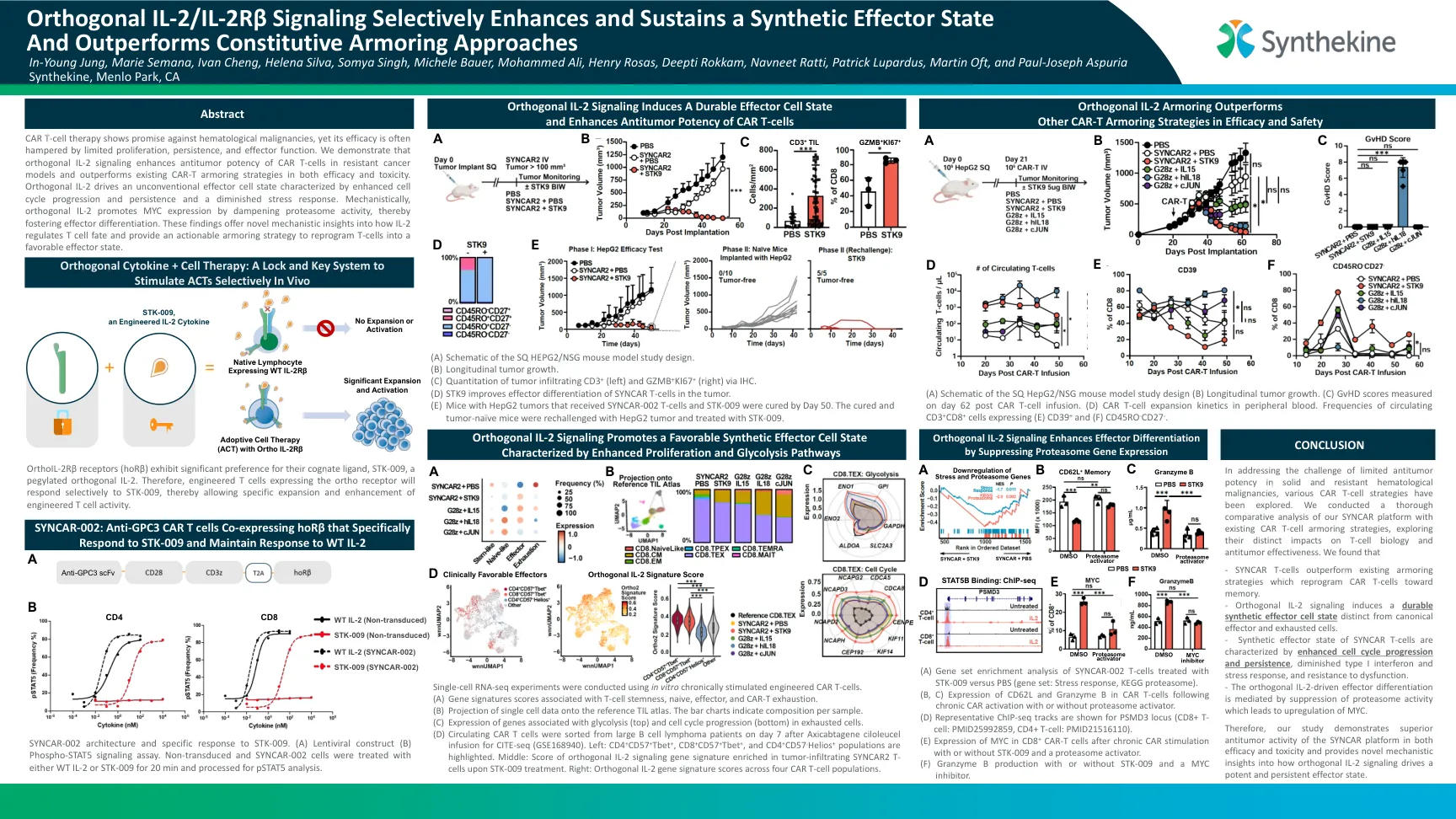
正交IL-2/IL-2Rβ信号有选择地增强和维持合成效应子状态,并且表现优于组成型装甲方法
CAR T细胞疗法表现出对血液学恶性肿瘤的有望,但其功效通常受到有限的增殖,持久性和效应子功能的阻碍。我们证明,正交IL-2信号传导在抗性癌症模型中增强了CAR T细胞的抗肿瘤效力,并且在功效和毒性中都胜过现有的CAR-T装甲策略。正交IL-2驱动非常规效应细胞态,其特征是细胞周期进展和持久性增强以及应力反应减少。 从机械上讲,正交IL-2通过抑制蛋白酶体活性促进MYC的表达,从而促进效应子分化。 这些发现提供了有关IL-2如何调节T细胞命运的新型机械见解,并提供了可行的装甲策略,以将T细胞重编程为有利的效应子状态。正交IL-2驱动非常规效应细胞态,其特征是细胞周期进展和持久性增强以及应力反应减少。从机械上讲,正交IL-2通过抑制蛋白酶体活性促进MYC的表达,从而促进效应子分化。这些发现提供了有关IL-2如何调节T细胞命运的新型机械见解,并提供了可行的装甲策略,以将T细胞重编程为有利的效应子状态。

使用相...
高位率无线通信要求高频率[1],例如24-GHz WLAN [2],IEEE802.11AD [3]和24-29 GHz 5G手机[4]。高频无线系统中的必需电路块之一是电压控制的振荡器(VCO)。进行正交信号处理[5,6,7,8,9]的[5,6,7,8,9] [10,11,12,12,12,13,14,15,16,17],但是,高频率VCO通常需要一个非常非常高的CMOS技术和/或特殊QMOS技术和特殊的QMOS技术阶段[18] 噪音。 因此,他们的过程成本可能很高。 在这封信中,提出了使用相调整架构来抑制相位噪声的正交VCO。 可以在不增加过程成本的情况下实现此体系结构。进行正交信号处理[5,6,7,8,9]的[5,6,7,8,9] [10,11,12,12,12,13,14,15,16,17],但是,高频率VCO通常需要一个非常非常高的CMOS技术和/或特殊QMOS技术和特殊的QMOS技术阶段[18] 噪音。因此,他们的过程成本可能很高。在这封信中,提出了使用相调整架构来抑制相位噪声的正交VCO。可以在不增加过程成本的情况下实现此体系结构。

一种正交激活的 CRISPR-Cas13d 纳米前药,可逆转化学耐药性,从而增强化学光动力疗法
通过运输氧化/还原形式的谷胱甘肽及其药物偶联物来改变细胞的氧化还原状态;并且与癌症的不良临床结果(例如预后不良)密切相关。4因此,MRP1 是耐药癌细胞的“致命弱点”之一。5越来越多的证据表明,通过基因沉默方法下调 MRP1 基因可以逆转 MRP1 介导的耐药性。6例如,已发现成簇的规律间隔的短回文重复相关蛋白 9 (CRISPR-Cas9) 技术可以逆转由 ATP 结合盒 (ABC) 转运蛋白介导的 MDR,由于其设计简单、靶区域灵活、编辑效率更高和多路复用,其结果明显高于其他基因编辑技术。 7 – 10 尽管取得了巨大进展,但大多数 CRISPR-Cas9 系统仍然存在一些棘手的问题,包括非靶标基因组改变和基因毒性、Cas9 特异性 T 细胞的潜在免疫风险以及不令人满意的靶向递送。8 为了应对这一挑战,RNA 引导的 VI 型 Cas 蛋白 CRISPR-Cas13d 已被证实可在不改变基因组的情况下敲低靶基因。11,12 重要的是,与 Cas9 蛋白相比,

Agilent AN 1298 通信系统中的数字调制
2.6 无线电发射机中的 I 和 Q I/Q 图特别有用,因为它们反映了使用 I/Q 调制器创建大多数数字通信信号的方式。在发射机中,I 和 Q 信号与相同的本地振荡器 (LO) 混合。90 度移相器放置在其中一个 LO 路径中。相隔 90 度的信号也称为彼此正交或正交。正交信号不会互相干扰。它们是信号的两个独立分量。重新组合时,它们将相加为复合输出信号。I 和 Q 中有两个独立信号,可以通过简单的电路发送和接收。这简化了数字无线电的设计。I/Q 调制的主要优点是能够轻松地将独立信号分量组合成单个复合信号,然后再将这种复合信号拆分成其独立分量。