XiaoMi-AI文件搜索系统
World File Search System活化

研究人员会开发模仿抗原的合成细胞,以改善T细胞活化| UCLA Health
天然细胞的粘弹性。微流体系统用于精确控制这些合成细胞的大小,形状和机械性能。它们的粘弹性使合成细胞可以更好地复制T细胞检测到的有机细胞相互作用,从而显着改善了T细胞激活,扩展和功能。


新的核法医学 - SIPRI
I. 对人体的辐射剂量 130 � II. 来自废弃地下核试验场的核素 131 � III.与全面禁核试条约核查有关的颗粒放射性核素 133 � 类别 1. 燃料材料的残留物 136 � 类别 2. 燃料材料的非裂变反应产物 139 � 类别 3. 裂变产物 141 � 类别 4. 非燃料弹材料的活化产物 142 � 类别 5. 地下爆炸周围的填塞(填充)材料和岩石中的活化产物 143 � 类别 6. 近地表大气爆炸下方地面中的活化产物 144 类别 7. 水下或近海面爆炸周围海水中的活化产物 144 � 类别 8. 大气爆炸周围空气中的活化产物 144 � 类别 9. 来自中子通量探测器的活化产物 145 � 类别 10. 添加的示踪剂 145 � 制定全面禁核试条约相关颗粒放射性核素最终清单核素 146 � IV. 与《全面禁试条约》国际监测系统有关的惰性气体放射性核素 153 � V. 与现场视察有关的颗粒和气体核素 154 �

AZD9291耐药非小细胞肺癌细胞来源的外泌体lnc-MZT2A-5:1诱导成纤维细胞活化
免疫治疗在部分晚期 NSCLC 患者中取得了很大进展(4)。靶向治疗和免疫治疗已取代传统的手术干预,改变了大多数患者的治疗进程,分子检测现已成为晚期肺腺癌患者的标准推荐(5)。奥希替尼 (AZD9291) 是第三代酪氨酸激酶抑制剂 (TKI),通过与突变 EGFR 的 C797 残基形成共价键,选择性地靶向激活 EGFR 突变和 T790M 抗性突变(6)。尽管 AZD9291 不仅在一线治疗中而且在 T790M 继发突变的挽救治疗中都取得了巨大成功,但不可避免地会出现获得性耐药,限制了其长期临床益处(7)。然而,AZD9291 的耐药机制仍未完全了解(8)。因此,进一步了解肺癌患者的耐药机制、进行靶向治疗及个性化治疗已成为该领域的研究热点。

CAGE 测序揭示了活化囊性纤维化巨噬细胞中 CFTR 依赖的 I 型 IFN 信号失调
强烈的、无法缓解的气道炎症反应会导致囊性纤维化 (CF) 患者的破坏性肺部疾病。巨噬细胞免疫功能失调可能是控制 CF 肺部疾病进展的一个关键方面,但其潜在机制尚不完全清楚。我们使用 5′ 端为中心的转录组测序来分析铜绿假单胞菌 LPS 激活的人类 CF 巨噬细胞,结果显示 CF 和非 CF 巨噬细胞在基线和激活后部署了截然不同的转录程序。这包括与健康对照相比,激活的患者细胞中 I 型 IFN 信号反应明显减弱,但在患者细胞中使用 CFTR 调节剂进行体外治疗以及通过 CRISPR-Cas9 基因编辑来纠正患者来源的 iPSC 巨噬细胞中的 F508del 突变后,这种反应是可逆的。这些发现表明,人类 CF 巨噬细胞中存在以前未被发现的免疫缺陷,这种缺陷依赖于 CFTR,并且可以通过 CFTR 调节剂逆转,从而为寻找 CF 中的有效抗炎干预措施提供了新的途径。
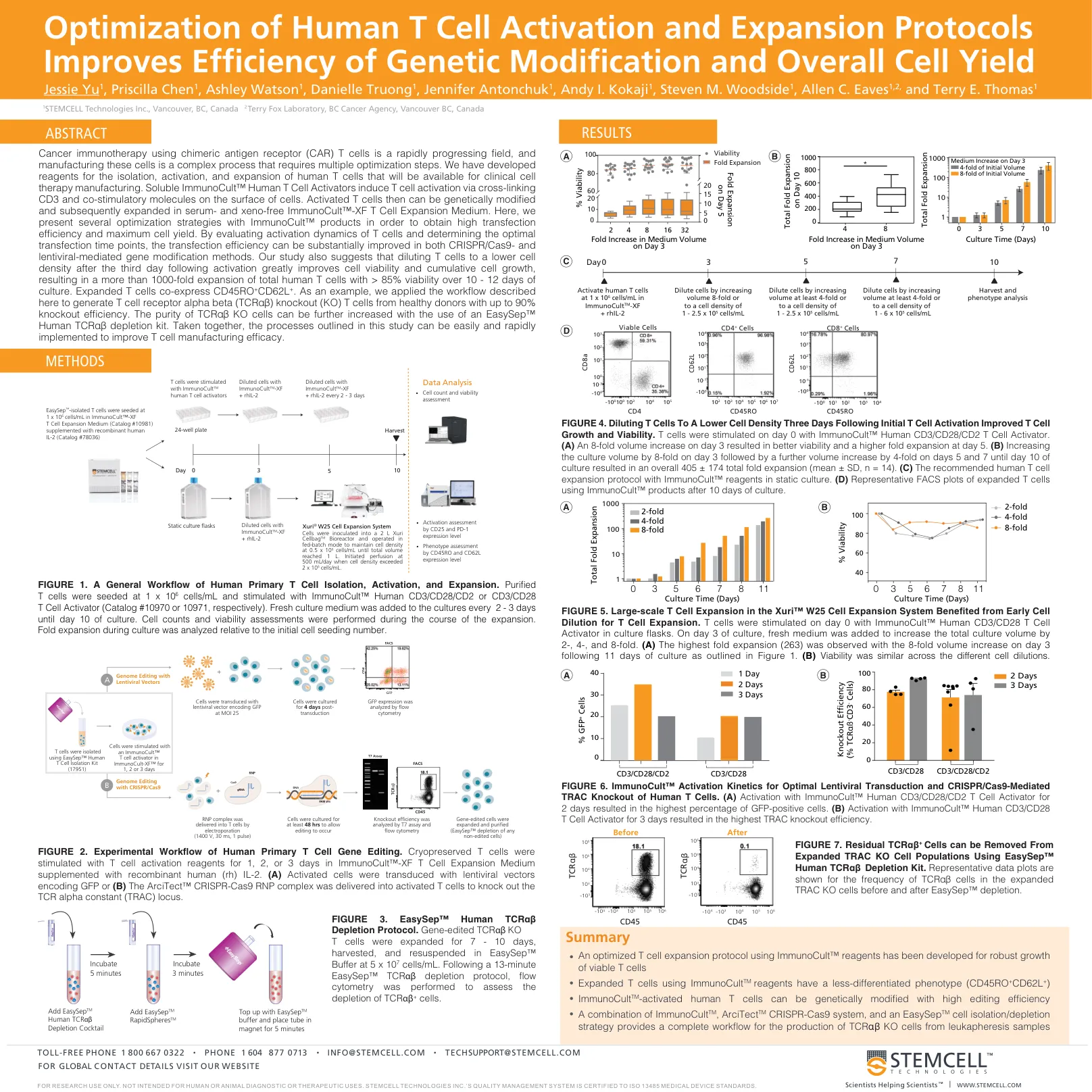
SP00235-优化人类T细胞活化和扩增方案可提高基因改造效率和总细胞产量
使用嵌合抗原受体 (CAR) T 细胞进行癌症免疫治疗是一个快速发展的领域,制造这些细胞是一个复杂的过程,需要多个优化步骤。我们已经开发出用于分离、激活和扩增人类 T 细胞的试剂,可用于临床细胞疗法制造。可溶性 ImmunoCult™ 人类 T 细胞激活剂通过交联细胞表面的 CD3 和共刺激分子来诱导 T 细胞活化。然后可以对活化的 T 细胞进行基因改造,随后在无血清和无异种的 ImmunoCult™-XF T 细胞扩增培养基中扩增。在这里,我们介绍了几种使用 ImmunoCult™ 产品的优化策略,以获得高转染效率和最大细胞产量。通过评估 T 细胞的活化动力学并确定最佳转染时间点,可以显著提高 CRISPR/Cas9 和慢病毒介导的基因修饰方法中的转染效率。我们的研究还表明,在激活后第三天将 T 细胞稀释至较低的细胞密度可大大提高细胞活力和累积细胞生长,从而使培养 10 - 12 天后的总人类 T 细胞扩增超过 1000 倍,活力 > 85%。扩增的 T 细胞共表达 CD45RO + CD62L + 。例如,我们应用此处描述的工作流程从健康供体中生成 T 细胞受体 α β (TCR αβ ) 敲除 (KO) T 细胞,敲除效率高达 90%。使用 EasySep™ 人 TCR αβ 耗竭试剂盒可以进一步提高 TCR αβ KO 细胞的纯度。总之,本研究中概述的流程可以轻松快速地实施,以提高 T 细胞制造效率。

免疫原性诱导化学疗法对黑色素瘤中免疫细胞活化和三级淋巴结构形成的影响
结果:单独的阿霉素,单独的吉西他滨和两种药物组合都放慢了肿瘤的生长,结合治疗表现出更明显的作用。与对照组相比,阿霉素组显示出更高的CD8 + T细胞和组织居民记忆T细胞(T RM)的纤维化,并且在CD8 + T亚群中的干扰素G,Granzyme B和穿孔蛋白的分泌增加以及B细胞和B细胞的活化。单独的阿霉素并与吉西他滨组合降低了调节性T细胞。此外,阿霉素治疗促进了HEV和TLS的形成。阿霉素治疗还上调了肿瘤细胞中CD8 + T细胞和程序性细胞死亡蛋白配体(PD-L)1中编程细胞死亡蛋白(PD)-1的表达。

通过经验价键模拟精确计算分支酸变位酶反应中的热力学活化参数
摘要:分支酸变位酶 (CM) 长期以来一直用作计算化学中基准测试新方法和工具的模型系统。尽管这些酶在文献中占有重要地位,但活化焓和熵在催化分支酸转化为预苯酸盐方面所起的作用程度仍有待商榷。了解这些参数是充分理解分支酸变位酶机制的关键。在本研究中,我们利用一系列温度下的 EVB/MD 自由能扰动计算,使我们能够从单功能枯草芽孢杆菌 CM 和铜绿假单胞菌的混杂酶异分支酸丙酮酸裂解酶催化的反应的活化自由能的阿伦尼乌斯图中提取活化焓和熵。与未催化反应相比,我们的结果表明,两种酶催化反应的活化焓均显著降低,而对活化熵的影响相对较小,表明酶催化的 CM 反应是焓驱动的。此外,我们观察到枯草芽孢杆菌的单功能 CM 比其混杂对应物更有效地催化此反应。过渡态反应途径的结构分析支持了这一点,从中我们确定了解释反应焓驱动性质以及两种酶之间效率差异的关键残基。

巨噬细胞中的基因筛选确定了 IFN γ 诱导的 MHCII 的新调节剂,这些调节剂有助于 T 细胞活化
摘要 细胞因子介导的宿主免疫激活是控制病原体的核心。干扰素-γ (IFN γ ) 是保护性免疫中的关键细胞因子,可诱导主要组织相容性复合体 II 类分子 (MHCII) 以扩增 CD4 + T 细胞活化和效应功能。尽管 IFN γ 诱导的 MHCII 起着核心作用,但其动态调节尚不明确。我们在小鼠巨噬细胞中使用全基因组 CRISPR-Cas9 筛选,确定了控制 MHCII 表面表达的基因。机制研究揭示了两条平行的 IFN γ 介导的 MHCII 控制途径,这两条途径需要多功能糖原合酶激酶 3 β (GSK3 β ) 或介导复合物亚基 16 (MED16)。这两种途径控制着 IFN γ 反应的不同方面,并且对于 IFN γ 介导的 MHCII 转录激活因子 Ciita 的诱导、MHCII 表达和 CD4 + T 细胞活化必不可少。我们的研究结果确定了之前未被重视的 MHCII 表达调节,这种调节对于控制 CD4 + T 细胞反应必不可少。