XiaoMi-AI文件搜索系统
World File Search System相关因子

使用 CRISPR/Cas9 进行 T 细胞 Nrf2/Keap1 基因编辑和实验性肾脏缺血-再灌注损伤
目的:T 细胞在肾脏缺血再灌注损伤 (IRI) 中发挥病理生理作用,核因子红细胞 2 相关因子 2/kelch 样 ECH 相关蛋白 1 (Nrf2/Keap1) 通路调节 T 细胞反应。我们假设成簇的规律间隔短回文重复序列 (CRISPR)/CRISPR 相关蛋白 9 (Cas9) 介导的 Keap1 敲除 (KO) 增强了 CD4+ T 细胞的 Nrf2 抗氧化潜力,而 Keap1 -KO CD4+ T 细胞免疫疗法可预防肾脏 IRI。结果:CD4+ T 细胞 Keap1-KO 导致 Nrf2 靶基因 NAD(P)H 醌脱氢酶 1、血红素加氧酶 1、谷氨酸-半胱氨酸连接酶催化亚基和谷氨酸-半胱氨酸连接酶修饰亚基显著增加。体外,Keap1-KO 细胞没有显示出衰竭迹象,在常氧条件下白细胞介素 2 (IL2) 和 IL6 水平显著降低,但在缺氧条件下干扰素 γ 水平升高。体内实验中,与接受未编辑对照 CD4+ T 细胞的小鼠相比,IRI 前过继转移 Keap1-KO CD4+ T 细胞可改善 T 细胞缺陷 nu/nu 小鼠的肾功能。与从对照肾脏中分离的未编辑 CD4+ T 细胞相比,IRI 后 24 小时从受体肾脏中分离的 Keap1-KO CD4+ T 细胞活性较低。创新:使用 CRISPR/Cas9 编辑小鼠 T 细胞中的 Nrf2/Keap1 通路是一种创新且有前景的免疫治疗方法,可用于治疗肾脏 IRI 以及其他实体器官 IRI。结论:CRISPR/Cas9 介导的 Keap1 -KO 增加了小鼠 CD4+ T 细胞中 Nrf2 调节的抗氧化基因表达,改变了对体外缺氧和体内肾脏 IRI 的反应。针对 T 细胞中 Nrf2/Keap1 通路的基因编辑是治疗免疫介导肾脏疾病的一种有前景的方法。抗氧化剂。氧化还原信号。38,959–973。

社论:炎症性疾病新型治疗分子和靶点的最新进展
炎症性疾病由多种以炎症为特征的疾病和病症组成,例如炎症性肠病、肝炎和类风湿性关节炎(Okin et al., 2012)。在炎症性疾病的病理条件下,免疫系统错误地攻击健康细胞或组织,导致慢性疼痛、发红、肿胀、僵硬和身体损伤(Marchetti et al., 2005)。炎症性疾病与多种潜在原因有关,包括饮食、压力和睡眠障碍。抗炎药物有助于预防或减少疾病进展。然而,常用药物经常伴有严重的不良反应。迫切需要开发新的炎症性疾病治疗方法并阐明关键基因和内在机制。诊断生物标志物在患者的诊断和治疗过程中的多个方面对疾病的治疗有用。炎症生物标志物多种多样,包括细胞因子/趋化因子、急性期蛋白、免疫相关效应物、活性氧和氮物质、前列腺素和环氧合酶相关因子、转录因子和生长因子 ( Brenner et al., 2014 )。Lin et al. 证明肠粘膜中的 lncRNA DLEU2 在肠道炎症时失调,可以作为溃疡性结肠炎的诊断生物标志物 ( Lin et al. )。他们将 DLEU2 鉴定为一种抗炎 lncRNA,通过负向调节 NF- κ B 信号通路来抑制肠道炎症 ( Lin et al. )。Huang et al.报道称,MHR(单核细胞与高密度脂蛋白的比率)和MAR(单核细胞与载脂蛋白A1的比率)是理想的促炎症标志物,可影响绝经后2型糖尿病女性骨微环境中由慢性炎症引起的骨稳态失衡(Huang et al.)。这些研究人员将生物标志物的研究课题扩展到炎症疾病。找到正确的治疗靶点是抗炎药物研发中最重要的方法。许多靶点负责抗炎作用,例如抑制细胞因子信号传导、降低白细胞活化、趋化性和募集。研究人员已经在这个研究课题中确定了几个靶点。K-Ras是一个研究得比较深入的致癌基因。Qi et al.报道称,抑制 K-Ras G13D 突变可通过 RAS/ERK 通路促进癌症干性和炎症 ( Qi et al. )。这一发现可能对理解 K-Ras G13D 突变对促进癌症干性和炎症的影响,在使用 K-Ras G13D 靶向疗法时具有重要意义

SGLT2抑制剂的心脏保护和抗癌作用
Akt¼蛋白激酶B; ALP¼碱性磷酸酶; a-sma¼a -smooth肌肉肌动蛋白; AMPK¼腺苷单磷酸 - 活化的蛋白激酶; ANP¼14钠肽; Arn¼血管紧张素受体Neprilysin抑制剂; AST¼天冬氨酸氨基转移酶; ATF-4¼激活转录因子4; BAX¼Bcl-2相关X蛋白; B-MHC¼B-肌球蛋白重链; bohb¼b-羟基丁酸酯; BNP¼B型纳特里尿肽; CAT¼过氧化氢酶; CFR¼冠状动脉储备; CK-MB¼肌酸激酶MB; CRS¼心脏综合征; CTNT¼心脏肌钙蛋白T;潮湿¼损伤相关的分子模式; dox¼阿霉素; ECG¼心电图; ef¼射血分数; EIF-2a¼真核生物起始因子2 a; Er¼内质网; ERK¼1.1.1/1/14; FGF¼FIMBLAST生长因子; FS¼部分缩短; g-csf¼1/1/14 GM-CSF¼1/1/1/14 GRP78¼葡萄糖调节的蛋白78; HTN¼高血压; I.P.¼腹膜内; IL¼白痴; IL¼白痴; IL¼白痴; iNOS¼诱导一氧化氮合酶; LDH¼14乳酸脱氢酶; LV¼左心室; lvedd¼左心室末端直径; lvesd¼左心室末端音直径; LVIDD¼左心内直径在末端末端;末端收缩处的LVIDS¼左心内直径; MDA¼MALONDIALLEDEDEDE; MMP¼基质金属肽酶; MPO¼髓过氧化物酶;雷帕霉素的mtor¼哺乳动物靶标; mybpc3¼结合蛋白C3; MyD88¼髓样差异反应88; NCD¼正常食物饮食; NF-kb¼核因子kappa-b; NLRP3¼NOD样受体蛋白3;无¼一氧化氮; NOX-1¼NADPH氧化酶1; NOX-2¼NADPH氧化酶2; NRF2¼核因子红细胞2 - 相关因子2; NT-Proanp¼n末端Pro - 心房纳地肽; NT-PROBNP¼N末端Pro - B型纳地尿肽; p38¼p38有丝分裂原激活的蛋白激酶; PARP¼聚(二磷酸腺苷 - 核糖)聚合酶; PERK¼蛋白激酶R样性内质网激酶; PGC¼过氧化物酶体增殖物 - 激活的受体共激活剂; PI3K¼磷酸肌醇3-激酶; PPAR¼过氧化物酶体增殖物 - 活化受体; QTC¼校正的QT; SIRT1¼SIRTUIN1; Sirt3¼Sirtuin3; Smad3¼母亲反对脱皮的同源物3; SOD¼超氧化物歧化酶; TGF¼转化生长因子; TLR9¼Toll样受体9; TNF¼肿瘤坏死因子; XO¼黄嘌呤氧化酶;其他缩写如表1所示。

引文 Igarashi A, Tsuji G, Murata R, Fukasawa S 和 Yamane S (2024), tapinarof 对日本患者皮肤屏障功能的改善作用
特应性皮炎 (AD) 是一种常见的慢性炎症性皮肤病 [1]。在许多国家,AD 的终生患病率估计为 15% 以上 [2]。持续剧烈瘙痒导致的明显皮肤病变和睡眠障碍显著影响 AD 患者的生活质量 [3,4]。AD 的发病机制是多因素的,涉及异常的免疫反应、遗传和环境因素以及皮肤屏障功能障碍 [5-7]。2 型辅助 T (Th2) 细胞因子,如白细胞介素 (IL)-4 和 IL-13,在 AD 的发展中起着重要作用 [2]。AD 的皮肤屏障功能障碍与角质层中一种关键的皮肤屏障相关蛋白 fi laggrin 基因的功能丧失突变有关 [8,9]。 Th2 细胞因子可下调皮肤屏障相关蛋白的表达,而 Th2 细胞因子在 AD 皮肤病变中过度表达,导致进一步的皮肤屏障功能障碍 [8,9]。与健康皮肤相比,AD 皮肤病变处的角质层水合 (SCH) 减少,经表皮失水 (TEWL) 增加 [10],这反映了 AD 患者皮肤水分含量较低是由屏障功能障碍导致的。AD 治疗的主要目标是维持长期缓解,即 AD 的症状和体征消失或极少,且不影响日常活动 [1]。外用皮质类固醇是目前治疗 AD 的主要手段;然而,长期使用外用皮质类固醇会导致特定的不良反应,如皮肤萎缩,并可能导致皮肤屏障功能障碍 [11]。尽管非甾体外用药物已经面世,但仍然需要能够长期使用并改善皮肤屏障功能的有效外用药物。他匹那洛夫是一种非甾体外用芳烃受体 (AhR) 激动剂 [12]。AhR 是一种胞浆配体依赖性转录因子。通过激活 AhR,他匹那洛夫上调皮肤屏障相关蛋白的表达,如 filagrin、hornerin 和 involucrin [12]。他匹那洛夫的药理作用还包括下调促炎性细胞因子表达和通过激活核因子红细胞 2 相关因子 2 通路上调抗氧化酶表达 [12,13]。因此,他匹那洛夫可以成为一种治疗 AD 的新型外用药物,其特征性药理作用包括改善皮肤屏障功能的潜力。迄今为止,已开展了多项临床研究,以评估他匹那洛夫在 AD 患者中的疗效和安全性 [ 14 – 17 ]。这些研究结果表明他匹那洛夫是有效的,并且具有可接受的安全性。在本研究中,我们试图评估他匹那洛夫对 AD 患者皮肤屏障功能的改善作用。

携带EGFR突变的小细胞肺癌患者
抽象背景二甲双胍(MET)是2型糖尿病的一线治疗方法,在治疗各种疾病(例如心血管疾病,神经退行性疾病,癌症和衰老)中起着有效的作用。然而,依赖MET的抗肿瘤免疫的潜在机制仍有待阐明。方法Mitotempo是线粒体超氧化物的清道夫,废除了MET的抗肿瘤作用,但没有消除抗编程细胞死亡(PD-1)抗体(AB)治疗。因此,我们研究了遇到的抗肿瘤效应的机制。葡萄糖转运蛋白(GLUT)-1,线粒体活性氧(MTROS),干扰素(IFN)-γ,Ki67,自噬标记,NF-E2相关因子2(NRF2)的激活标记和乳腺脂蛋白复合物1(Mammaycin Confictrc 1(Mimamalian Infilc)1(mmAMAMAYCINC)1(MIMMAYCIN COFFILC 1)通过流式细胞仪分析检查淋巴细胞(CD8TIT)。此外,还使用有条件的NRF2和p62小鼠来检测这些标记,并监测体内肿瘤生长。对CD8TIT和肿瘤细胞进行 RNA测序。 黑色素瘤细胞含有IFN-γ受体(IFNγR)细胞质域缺失突变体过表达,并用于使用Seahorse Flux分析仪来表征这些肿瘤细胞的代谢谱。 结果MET给药可提高MTROS和细胞表面Glut-1,从而导致CD8TIT中的IFN-γ产生。 mtros以糖酵解依赖性方式激活NRF2,从而诱导自噬,谷氨酰胺溶解,mTORC1和p62/sqSTM1的激活。RNA测序。黑色素瘤细胞含有IFN-γ受体(IFNγR)细胞质域缺失突变体过表达,并用于使用Seahorse Flux分析仪来表征这些肿瘤细胞的代谢谱。结果MET给药可提高MTROS和细胞表面Glut-1,从而导致CD8TIT中的IFN-γ产生。mtros以糖酵解依赖性方式激活NRF2,从而诱导自噬,谷氨酰胺溶解,mTORC1和p62/sqSTM1的激活。在丝氨酸351(p-p62(S351))上,p62的MTORC1依赖性磷酸化也参与了NRF2的激活。CD8TIT中NRF2的条件缺失消除了MTORC1激活和抗肿瘤免疫。 与抗PD-1 AB的作用协同作用,满足CD8TIT增殖和IFN-γ分泌,从而导致肿瘤细胞中糖酵解和氧化磷酸化的降低。 因此,在CD8TIT中,GLUT-1以及活化的树突状细胞的扩展升高。 此外,缺乏IFNγR信号的肿瘤细胞消除了CD8TIT的IFN-γ的产生和增殖。CD8TIT中NRF2的条件缺失消除了MTORC1激活和抗肿瘤免疫。与抗PD-1 AB的作用协同作用,满足CD8TIT增殖和IFN-γ分泌,从而导致肿瘤细胞中糖酵解和氧化磷酸化的降低。因此,在CD8TIT中,GLUT-1以及活化的树突状细胞的扩展升高。此外,缺乏IFNγR信号的肿瘤细胞消除了CD8TIT的IFN-γ的产生和增殖。

胰腺β单元格:社论
胰腺β细胞通过作为胰岛素的主要来源来维持葡萄糖稳态方面起着关键作用。这些细胞负责胰岛素的合成,储存和释放,该胰岛素的合成,储存和释放是由于身体代谢状态的变化而受到严格调节。由于β细胞在糖尿病病理生理学中的核心作用,其生物学引起了科学界的显着兴趣。更好地了解β细胞生物学的多个方面可能会导致制定新的预防策略和治疗,从而延迟或停止疾病进展。这个特刊“胰腺β细胞”介绍了14篇文章的集合,其中包括五篇原始论文和9篇评论,突出了Beta细胞研究的各个方面。此问题的重点是控制β细胞质量膨胀和存活的分子机制,尤其强调了成熟β细胞功能的显着途径。贡献涵盖了广泛的主题,包括氧化应激对β细胞的影响[1-3],组织间通信[4-7],以及β细胞质量和功能的主调节剂[8-10]等。Mukai及其同事[1]回顾了氧化应激和β细胞抗氧化剂机制的作用,总结了β细胞中抗氧化酶的低表达和氧化应激如何损害胰岛素的分泌。作者建议核因子2与2相关因子2(NRF2)是β细胞抗氧化剂反应的主要调节剂。作者建议TMAO可能在糖尿病生成条件下介导一种补偿性的蛋白质作用。Wu及其同事[3]探索了在氧化应激条件下4-辛基伊替酸(4-OI)对胰腺β细胞的影响。研究人员发现,4-OI治疗减少了活性氧的产生,抑制细胞死亡途径激活和炎性细胞因子分泌,并逆转了缺氧诱导的细胞死亡,这表明4-OI可能在氧化应激条件下增强β细胞存活。此外,此问题还包括有关分子和其他组织中产生因素对β细胞功能的影响的文章。Krueger及其同事[4]研究了肠道微生物代谢产物三甲胺N-氧化物(TMAO)对功能性β细胞质量的影响。研究人员发现,尽管据报道患者的2型糖尿病(T2D)水平升高,但TMAO保护了β细胞功能并改善氧化和内质网应激。Fernandez-Millan及其同事[5]讨论了诸如T2D等代谢疾病病理学中的组织间交流的重要性,强调了如何理解Beta细胞与代谢和非代谢组织进行通信的方式提供了新的研究领域。他们强调了来自各种器官和组织对β细胞生物学的分泌因素的影响,这表明血时间交流可以为糖尿病研究提供新的机会。内分泌胰岛与胰腺中外分泌细胞的物理接近允许这些相邻细胞类型之间的旁分泌相互作用。作者强调了在这些情况下对糖尿病进行早期诊断的重要性。外分泌疾病对β细胞的影响是Ciochina and Floeagues的评论[6],它描述了慢性胰腺炎,急性胰腺炎,囊肿性肿瘤,胰腺癌,胰腺癌,胰腺癌,胰腺切除术以及Autoimmmune Pancreatiation and Autoimmune Pancreatiation如何影响Beta Celle和Diabect。Kryvalap及其同事[7]回顾了在外分泌胰腺中表达的蛋白酶和Serpin蛋白酶抑制剂对胰岛病理生理学的影响。作者探讨了对抑制或增强蛋白酶的反对意见

Ankrd11 是一种染色质调节剂和 KBG 综合征风险基因,是心脏神经嵴细胞生物学和心脏发育的关键调节剂
57 先天性心脏缺陷 (CHD) 是最常见的出生缺陷类型,全球发病率为 58 1% (Wu et al., 2020)。CHD 中很大一部分是圆锥动脉干缺陷,59 这是由于心脏流出道 (OFT) 重塑不当引起的,OFT 是连接心室和咽弓动脉 (PAA) 的胚胎结构 (Keyte and 61 Hutson, 2012; Neeb et al., 2013)。在胚胎发生过程中,OFT 被重塑为成熟的主动脉和肺动脉,以分离含氧血液和缺氧血液的循环。63 该过程由胚胎心脏的不同组织之间的相互作用协调:64 第二心脏场衍生细胞、OFT 腔内心内膜细胞和心脏神经嵴细胞 (CNCC)。 CNCC 功能失调是许多圆锥动脉干缺陷的主要原因 66 ,也是许多已知的多系统发育障碍的诱因(Neeb 等人,2013;Vega-67 Lopez 等人,2018)。68 CNCC 起源于神经管边界,并迁移到 PAA 和 OFT,在那里 69 它们促进血管重塑并形成动脉平滑肌细胞内壁 70 (Keyte and Hutson,2012)。填充 OFT 垫的 CNCC 融合在一起形成 71 主动脉肺动脉 (AP) 隔,该隔从 OFT 的远端边缘发展直至与室间隔融合 72,形成主动脉和肺动脉两条不同的血管。 73 在小鼠中,OFT 分隔发生在胚胎第 11.5 天 (E) 和第 13.5 天之间 (Krishnan 等人,74 2014)。75 神经嵴发育需要复杂的基因表达时空调控。76 大多数研究集中在信号通路和转录因子上,而 77 表观遗传调控的研究相对不足 (Martik and Bronner, 2017; Neeb 等人,78 2013; Stefanovic 等人,2021; Yan 等人,2021)。尽管如此,已发现一些表观遗传调节因子对 CNCC 的正常发育至关重要,这些调节因子与各种先天性疾病有关,包括 Coffin-Siris (Brg1)、CHARGE (CHD7) 和 Williams (BAZ1B) 综合征 (Barnett 等人,2012 年;Li 等人,2013 年;Yan 等人,2021 年;Yan 等人,82 2020 年)。83 染色质调节因子对于基因表达的时空调控至关重要,这对于协调复杂的 OFT 重塑过程必不可少,尽管神经嵴辅助心脏发育中的大部分过程仍不清楚 (Yan 等人,2021 年)。 ANKRD11 86(锚蛋白重复域 11;以前称为 ANCO1)是一种染色质调节剂,可募集 87 组蛋白乙酰化修饰蛋白,例如组蛋白去乙酰化酶 HDAC3 和 P/CAF 88(p300/CBP 相关因子)乙酰转移酶复合物亚基,以调节全局基因 89 表达(Gallagher 等人,2015 年;Li 等人,2008 年;Zhang 等人,2004 年)。90 ANKRD11 或含有 ANKRD11 的 16q24.3 微缺失中的杂合变异会导致 KBG 综合征(OMIM 91 #148050),一种常染色体显性多系统发育障碍。患有 KBG 92 综合征的患者表现出整体发育迟缓、身材矮小、颅面缺陷和智力 93 障碍(Digilio 等人,2021 年;Gnazzo 等人,2020 年;Goldenberg 等人,2016 年;Handrigan 等人,94 2013 年;Low 等人,2016 年;Murray 等人,2017 年;Ockeloen 等人,2015 年;Sirmaci 等人,2011 年;95 Willemsen 等人,2010 年)。约 40% 的患者有心血管缺陷,包括 96 主动脉缩窄、动脉导管未闭、瓣膜狭窄和室间隔缺损 (VSD) 97 (Digilio 等人,2021;Guo 等人,2022;Kierzkowska 等人,2023)。值得注意的是,这些心血管 98 缺陷表明 CNCC 功能可能失调 (Neeb 等人,2013;Vega-Lopez 等人,2018)。然而,Ankrd11 在 CNCC 命运或心脏发育中的作用尚不清楚。100
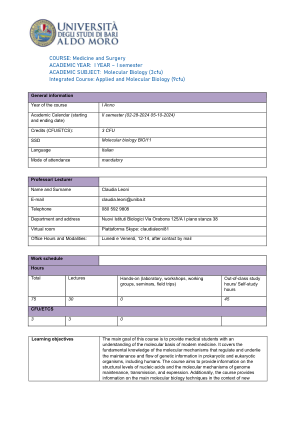
分子生物学(3cfu)综合课程
超螺旋和拓扑性质。拓扑异构酶。细菌类核。组蛋白和核小体的性质和组装。染色质的高级结构。组蛋白的翻译后修饰。溴多胺和染色质结构域。表观遗传学。原核生物和真核生物的基因组。复制模型。DNA合成。细菌DNA聚合酶。校对和缺口翻译。复制子模型。OriC和半甲基化。Ter/Tus。真核细胞核中的复制工厂。ARS结构和复制控制。酶学。前RC和前启动复合物。复制抑制剂,如化疗药物和抗病毒药物。端粒和端粒酶的结构、功能和意义。DNA损伤和修复。基因组作为动态实体。体细胞和种系突变。SNP。内在和外在损伤。化学和物理诱变剂。原核生物和真核生物中的去除、逆转和损伤避免系统。MUT 系统。BER 系统。糖基化酶的重要性。安全系统。NER 系统:UvrABCD 和 XP 蛋白。GG-NER 和 TC-NER。光解作用、MGMT、AlkBH。损伤耐受机制。TLS。细菌中的 SOS 反应。单丝和双丝断裂。HR 和 NHEJ。由于修复系统突变而导致的人类疾病。位点特异性重组。重组酶。Lambda 噬菌体。Cre-Lox 系统和 KO 小鼠。简单和复杂的转座子。SINE 和 LINE 元素、Alu 序列。原核生物和真核生物中的 RNA。结构、类型和特性。细菌 RNA 聚合酶和相关因子。转录单位。转录步骤。细菌启动子中的共识序列。终止机制。抑制剂。 Lac、ara 和 trp 操纵子。阳性和阴性对照。真核细胞中的 RNA 类别。RNA 聚合酶 (CTD) 的结构和功能。三种启动子的特征。基础转录机制。TFIIH。反式激活因子、辅激活因子。CpG 岛甲基化。组蛋白密码。长程调节剂。DNA 结合蛋白的功能域 (HTH、HD、HLH、ZF、LZ)。RNA 成熟、核运输和转录后控制。加帽类型。添加 polyA。CTD 的变化。外显子和内含子。外显子改组。四类内含子及其去除机制。剪接体和剪接位点。AT-AC 剪接。EJC 复合体。可变剪接。ESE 和 ESS 序列、SR 和 hnRNP 蛋白。SMN 基因。剪接和病理。rRNA 和 tRNA 加工反应。核糖体基因。 SnoRNA 和核仁功能。RNA 编辑。插入和转换编辑。人类 RNA 编辑的示例。细胞核和细胞质中的 RNA 周转。外泌体。无义介导的 mRNA 衰变 (NMD)。非编码 RNA。小 RNA 在细胞中的功能。RNA 干扰。siRNA。微小 RNA 的生物发生。miRNA、长链非编码 RNA、环状 RNA 的作用机制。逆转录病毒的一般信息。遗传密码和翻译。遗传密码的性质和特征。线粒体密码。ORF。tRNA 的特征。不常见碱基。aa-tRNA 合成酶的功能和类别。遗传密码的翻译重编码和扩展。SeCys。核糖体是一种核酶。原核生物和真核生物的翻译阶段。不同的启动机制。能量成本。NSMD。细菌中的 tmRNA。抑制剂。蛋白质的翻译后修饰、分选和降解。折叠和错误折叠。朊病毒。HSP60 和 HSP70。泛素和泛素化系统。SUMO 化糖基化。蛋白酶体。肽信号。蛋白质分选。线粒体输入。线粒体基因组细胞中的线粒体可塑性。人类线粒体基因组。遗传、结构、复制及其表达的原理。线粒体 DNA 中的改变。DNA 克隆的原理。修饰限制系统。克隆载体。cDNA 合成。基因组 DNA 和 cDNA 文库。TA 克隆。表达克隆。基因表达沉默。基因治疗。数据库。基因组编辑元件(Talen、Zn 指、CRISPR/Cas9 系统)。PCR 和 DNA 测序。PCR 的特性。PCR-RFLP。实时 PCR、DNA 测序。NGS。核酸杂交。杂交原理。熔点和严格性。探针制备:切口平移。Southern、Northern、杂交测定。蛋白质印迹。

铁凋亡和杯凋亡
缩写:5-FU,5-氟尿嘧啶;AA-CoA,花生四烯酸辅酶 A;ABCC1,ATP 结合盒,C 亚家族(CFTR/MRP),成员 1;ACC,无定形碳酸钙;ACLS4,酰基辅酶 A 合成酶家族 4;AdA-CoA,肾上腺酸辅酶 A;ALDH,醛脱氢酶;AML,急性髓细胞白血病;APC,抗原处理细胞;ARE,抗氧化反应元件;ART,青蒿素;BAX,BCL-2 相关 X 蛋白;BCL-2,B 细胞淋巴瘤 2;BTIC,脑肿瘤起始细胞;CBR,临床受益率;CLL,慢性淋巴细胞白血病;CNSI-Fe(II),碳纳米颗粒负载铁;CQ,氯喹;CRPC,去势抵抗性前列腺癌; CSC,癌症干细胞;CTL,细胞毒性 T 淋巴细胞;CuET,二乙基二硫代氨基甲酸铜 (II);DAMP,损伤相关分子模式;DFO,去铁胺;DHA,双氢青蒿素;DLAT,丙酮酸二氢硫酰赖氨酸残基乙酰转移酶成分;DMT1,二价金属转运蛋白 1;DOX,阿霉素;DRD2,多巴胺 D2 受体;DSF,双硫仑;EGFR,表皮生长因子受体;EMT,上皮-间质转化;ER,内质网;ETO,依托泊苷;FDX1,铁氧还蛋白 1;FER-1,铁抑制蛋白 1;FMN,基于框架的纳米剂;FPN1,铁转运蛋白 1;FTH1,铁蛋白重链 1; FTL1,铁蛋白轻链 1;GPX4,谷胱甘肽过氧化物酶 4;GSH,谷胱甘肽;GSS,谷胱甘肽合成酶;H 2 O 2,过氧化氢;HNC,头颈癌;HO-1,血红素加氧酶-1;ICD,免疫细胞死亡;ICIs,免疫检查点抑制剂;IDH1,异柠檬酸脱氢酶 1;IFN-γ,干扰素-γ;IREB2,铁反应元件结合蛋白 2;IREs,铁反应元件;IRP-2,铁调节蛋白 2;IRPs,铁调节蛋白;JAK,Janus 酪氨酸激酶;KEAP1,kelch 样 ECH 相关蛋白 1;KRAS,Kirsten 大鼠肉瘤病毒致癌基因同源物;LA,硫辛酸; LC3II,微管相关蛋白 1 轻链 3α;LDH,乳酸脱氢酶;LiMOFs,锂基金属有机骨架;LIPRO-1,利普司他丁 1;LOX,脂氧合酶;LPCAT3,溶血磷脂酰胆碱酰基转移酶 3;MDA,丙二醛;MFC-Gem,载吉西他滨的碳质纳米粒子;MGMT,甲基鸟嘌呤甲基转移酶;MMNPs,磁性介孔二氧化硅纳米粒子;MMP-2,金属蛋白酶-2;MnFe 2 O 4 ,锰铁氧体;mRNAs,信使 RNA;NEPC,神经内分泌前列腺癌;NF- κ B,活化 B 细胞的核因子 κ 轻链增强子;NFS1,半胱氨酸脱硫酶;NK,自然杀伤细胞; NOX,NADPH 氧化酶 1;NRF2,核因子红细胞 2 相关因子 2;NSCLC,非小细胞肺癌;OC1,耳蜗毛细胞;OS,总生存率;P62,隔离小体 1;PET,正电子发射断层扫描;P-GP,P-糖蛋白;PCC,持久癌细胞;PCN(Fe) MOFs,Fe 3 + 卟啉金属有机骨架上的 PEG;PD-L1,程序性死亡配体 1;PDAC,胰腺导管腺癌;PEG,聚乙二醇;PGE2,前列腺素 E2;PGRMC1,孕酮受体膜成分 1;PHPM,ROS 敏感聚合物;PTX,紫杉醇;PUFA,多不饱和脂肪酸;PUFA-OOH,磷脂多不饱和脂肪酸过氧化物;RIPK-1/2/3,受体相互作用丝氨酸/苏氨酸蛋白激酶 1/2/3;ROS,活性氧;RR,反应率;siRNA,小干扰 RNA;siSLC7A11,SLC7A11 siRNA;SLC3A2,溶质载体家族 3 成员 2;SLC40A1,溶质载体家族 40 成员 1;SLC7A11,溶质载体家族 7 成员 11;STAT1,信号转导和转录激活因子 1;TAM,肿瘤相关巨噬细胞;TCA,三羧酸循环;TFR,转铁蛋白受体;TME,肿瘤微环境; TMZ,替莫唑胺;TP53,细胞肿瘤抗原 p53;TRADD,肿瘤坏死因子受体 1 型相关死亡结构域蛋白;TTP,进展时间;US FDA,美国食品药品管理局;UTRs,非翻译区;VDAC,电压依赖性阴离子通道;xCT,谷氨酸-胱氨酸反向转运蛋白;Z-VAD-FMK,羧苄氧缬氨酰丙氨酰天冬氨酰-[O-甲基]-氟甲基酮;γ-GCS,γ-谷氨酰半胱氨酸合成酶。 * 通讯作者。电子邮箱地址:mateusz.kciuk@biol.uni.lodz.pl (M. Kciuk)。