XiaoMi-AI文件搜索系统
World File Search System红细胞

文章使用镰状细胞病和健康供体诱导的多能干细胞对 2D 和 3D 红细胞分化方案进行评估
摘要:背景:镰状细胞病 (SCD) 是一种由 HBB 基因点突变引起的高度流行的遗传性疾病,可导致慢性溶血性贫血和血管闭塞事件。患者来源的诱导性多能干细胞 (iPSC) 有望为开发具有抗镰状细胞活性的药物筛选新预测方法带来希望。在本研究中,我们评估并比较了使用健康对照和 SCD-iPSC 的 2D 和 3D 红细胞分化方案的效率。方法:对 iPSC 进行造血祖细胞 (HSPC) 诱导、红细胞祖细胞诱导和终末红细胞成熟。通过流式细胞术分析、菌落形成单位 (CFU) 测定、形态学分析和基于 qPCR 的 HBB 和 HBG2 基因表达分析来确认分化效率。结果:2D 和 3D 分化方案均诱导了 CD34 + /CD43 + HSPC。3D 方案对 HSPC 诱导表现出良好的效率 (>50%) 和高生产率 (45 倍),并增加了 BFU-E、CFU-E、CFU-GM 和 CFU-GEMM 集落的频率。我们还产生了 CD71 + /CD235a + 细胞 (>65%),与 3D 方案开始时相比,细胞扩增了 630 倍。红细胞成熟后,我们观察到 95% 的 CD235a + /DRAQ5- 去核细胞、正染色性红细胞,以及与成人 HBB 相比胎儿 HBG2 表达增加。结论:使用 SCD-iPSC 和比较分析确定了一种用于红细胞分化的稳健 3D 方案;然而,成熟步骤仍然具有挑战性,需要进一步开发。
社论
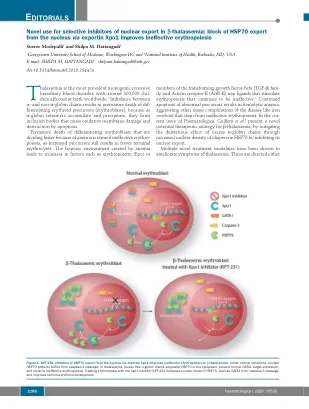
改善未结合的A -Globin和非A -Globin链之间的平衡或纠正无效的红细胞。修饰的TFG-β家族受体拮抗剂,如Sotatercemp(ACE-011)和Luspatercept(ACE-536)阻止配体与ACTR-II受体结合,并随后激活SMAD4信号通路,4改善Ery-Throid Throid Cell和红细胞的产生。通过CRISPR Therapeatics成功的基因疗法实现了未结合A -Globin与非A -Globin链的异常比例,并得到了波士顿顶点药物的支持。称为CTX001的体细胞疗法使用了编辑的患者自己的造血干细胞(HSC)来刺激胎儿血红蛋白的产生。5通过XPO1抑制HSP70的细胞内局部局部局部局部可能会合并这两个治疗目标。几条证据表明,红细胞使用分子伴侣在红细胞发育过程中对不稳定的过量A -Glo- bin链分割,6-8,因此,靶向这种伴侣的靶向时,当过量的globobin tetramer会累积时,靶向这种伴侣可能在β -tha -thaplamasemia中有用。许多组指出,分子伴侣HSP70在红细胞9-11中积聚至高水平,对于简化胚芽成熟很重要。11正常的人红细胞成熟需要在成熟后期的caspase-3瞬时激活,以防止过度的红细胞生产。激活的胱天蛋白酶可以切割GATA-1,从而导致成熟停滞和/或凋亡。12 Ribeil等。 14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。12 Ribeil等。14 GATA-1不再受到保护,导致末期成熟停滞和凋亡。表明EPO会导致HSP70转移到核中,结合GATA-1并保护其免受caspase-3裂解。相反,在EPO剥夺期间,HSP70被排除在细胞核中,而GATA-1被cas-pase-3裂解,导致凋亡死亡。13因此,HSP70的细胞内位置的改变似乎在红细胞生存力中起关键作用(图1)。在β-丘脑贫血中观察到的无效性红细胞生成的特征是在多染色体阶段加速了红细胞分化,成熟停滞和凋亡。在人β-thal虫蛋白粒细胞的成熟过程中,HSP70直接通过过量的游离A-格珠蛋白链直接在细胞质中螯合(图1)。核定靶向的HSP70突变体或caspase-3-无分解的GATA-1突变体恢复了β-thal核阿无血成红细胞的终末成熟。14在Haematologica,Guillem等。3跟进这种机制,以表明导出蛋白1(XPO1)调节在正常条件下HSP70在红细胞中的HSP70的核质质位置。Guillem等。证实,用XPO1抑制剂KPT-251治疗红细胞增加了HSP70的核水平,从caspase-3裂解中救出了GATA1,并改善了末端红细胞原理(图1)。尽管使用核出口的选择性抑制剂(SINE)用于治疗淋巴瘤和多骨髓瘤

医疗政策 - 镰状细胞病的基因疗法
功能失调的血红蛋白 S 产生的血红蛋白会在患者的红细胞中形成聚合物。在健康个体中,红细胞柔韧而圆润,因此可以轻松穿过血管。患有镰状细胞病的红细胞呈镰状或新月状,导致血液流经血管时速度减慢或堵塞。这会导致血管阻塞和缺血;红细胞寿命缩短,导致血管内和血管外溶血,粘稠的红细胞表面会增加对血管内皮的粘附,从而导致血管阻塞并导致血管增生性病变。2、复发性急性疼痛危机或血管闭塞危机是镰状细胞病最常见的表现。3、患者还会出现急性并发症,包括严重感染和非感染性并发症,如中风、肾坏死和异常勃起。 4,急性胸部综合征是一种可能危及生命的并发症,可能包括胸痛和呼吸急促等症状。5,慢性并发症可出现在多个器官中,包括青春期延迟、缺血性坏死、皮肤溃疡、慢性疼痛、神经认知障碍、慢性肾损伤、肺动脉高压、心血管疾病,并可能导致早期死亡。4,

2025 年 2 月 3 日 DHDTC DAL - NY.gov
医疗保健提供者依赖独立的社区供应商提供的血液制品(例如全血、血液成分(包括血浆)、新鲜冷冻血浆、采血后 24 小时内冷冻的血浆、红细胞、洗涤红细胞、白细胞减少红细胞、粒细胞和冷沉淀以及白蛋白和免疫球蛋白等衍生物)。假期和近期事件导致血液制品供应暂时减少,并造成血液供应紧张,包括这些产品的主要供应商的血液供应紧张。这给维持这些基本用品的供应带来了挑战。供应商必须采取积极主动的策略来节约血液供应库存,并确保有效和高效地利用现有库存。

509。骨髓衰竭和癌症易感综合征:先天性:海报II
HSP70伴侣GATA1,关键红细胞转录因子。没有HSP70,GATA1很容易通过caspase3。没有GATA1,关键红系分化靶标的转录未被激活,导致细胞凋亡和红细胞分化的延迟。3先前在RPL突变模型中已经报道了HSP70在DBA中的作用。4

腺苷信号传导抑制红细胞生成并促进髓样分化
造血是由骨髓中造血干细胞(HSC)产生所有血细胞的过程。促红细胞生成和颗粒状是造血的两个主要分支,分别是红细胞(RBC)和中性粒细胞的生产。虽然红细胞和髓样分化均来自相同的常见髓样祖细胞(CMP),但这两个过程之间的相互作用是复杂的,并且由不同的内在和外在因素紧密地策划,这些因子调节了祖细胞对一个细胞谱系或另一个细胞谱系或另一个细胞谱系的组合。1个末端红细胞生成和粒状植物发生在红细胞岛上,这些岛屿是骨髓中的专门微环体,该微晶体由中央宏观噬菌体组成,周围环绕着红细胞和中性粒细胞前体。2这些结构构成了独特的细胞微环境,并且通过提供必需的营养素,去除细胞碎片以及分泌细胞因子和生长方面来支持细胞增殖和分化至关重要。3越来越多的证据表明,在这些壁ches中发生平衡的微环境提示以及代谢物的运输和信号,还有其他

路易斯安那州镰状细胞委员会立法报告
镰状细胞病 (SCD) 是一种遗传性疾病,其特征是红细胞中一种名为血红蛋白的蛋白质。红细胞负责将血液中的氧气输送到身体各组织。正常的红细胞含有血红蛋白 A,呈圆形。然而,血红蛋白 S 的存在会导致红细胞变成镰状,这种形状的变化会导致红细胞阻塞血管,阻碍氧气输送。其后果可能是剧烈疼痛(也称为镰状细胞危象),并可能导致脑、肝、肾、骨骼和脾脏的永久性损伤。因此,SCD 患者感染、中风、心脏病和肾衰竭的风险增加。此外,SCD 的体征和症状因人而异,疼痛发作往往毫无征兆。症状的复杂性和多变性需要专业的临床指导和高级护理协调,包括咨询和教育。过去,SCD 被认为是“儿童疾病”,很少有患者能活到成年。 1 然而,医疗改革的努力已经大大提高了 SCD 的识别率,从而促进了治疗的进步,并在拥有适当资源的情况下显著提高了患者到 60 多岁的预期寿命。2 根据路易斯安那州新生儿筛查计划的数据,2023 财年有 73 名婴儿出生时患有血红蛋白疾病。其中,在此期间有 56 名新生儿被诊断出患有最严重的变异血红蛋白 S。此外,2023 财年有 2,385 名婴儿出生时为血红蛋白疾病携带者。大多数被确诊为携带者的婴儿(n=1,744)被诊断为镰状细胞 S 性状。其余被确诊为携带者的婴儿包括 527 名

镰状细胞病:基因治疗
要了解基因疗法,您需要了解红细胞和镰状细胞病。 • 红细胞将氧气从肺部输送到身体的所有组织。它通过一种称为血红蛋白的蛋白质来实现这一点。一种称为β珠蛋白基因的基因控制着血红蛋白过程。 • 镰状细胞病患者的β珠蛋白基因存在缺陷(破损)。缺陷基因导致红细胞呈新月形。它们粘在一起并阻塞小血管中的血流。这会导致疼痛和其他并发症。 • 基因疗法添加或用正常工作的基因替换有缺陷的基因。镰状细胞病有两种类型的基因疗法: − 基因添加使用载体或载体将基因的新拷贝放入细胞中。 − 基因编辑由科学家完成。他们在有缺陷的 DNA 处切割并替换

马等离子体中RHEPO/DPO的检测和定量
重组人红细胞生蛋白(RHEPO)2和darbepoetin-alpha(DPO)3是通过刺激红细胞生产来治疗贫血的基于基因工程的蛋白质药物。这些代理人刺激红细胞生产的能力导致了人类和马运动员的使用和滥用,因此违反了公平竞争的规则,导致其分类为赛马业被禁止的物质。此外,继续对马的行政管理可能导致贫血。3尽管马的Rhepo对马的负面方面,但由于浓度非常低,因此难以识别和合法的识别和合法性的方法来识别和确认Rhepo/DPO的方法。样本收集通常仅在竞争后才获得,该样本在管理后可能超过72小时。RHEPO/DPO的测试进一步与通常发现药物的基质的复杂性 - 血浆和尿液相混淆。

2024指南
可遇到另外的细胞减少症(中性粒细胞减少症比血小板减少症的频率更高),婴儿的短暂性血小板病。2除了患有GATA1突变的病例。3强烈暗示DBA综合征;但是不够具体,无法进行诊断。仅在专业实验室中进行4个研究测试;在模棱两可或非信息遗传学的情况下有用。 5通常在成年人中呈现。 6这些IBMF通常表现出多型细胞减少症,并且经常出现其他影响多器官系统的疾病特异性异常。 这种区别特征可以帮助将这些条件与DBA综合征区分开,该疾病最初以孤立的红细胞发育不全表现出来。 缩写:BM;骨髓; EADA,红细胞腺苷脱氨酶; HBF,胎儿血红蛋白; SLE,全身性红斑狼疮; prca,纯红细胞性植物; CLL,慢性淋巴细胞性白血病; LGL,大颗粒状淋巴细胞性白血病; CT,计算机断层扫描; MRI,磁共振成像; IBMF,继承的骨髓衰竭综合征; FA,Fanconi贫血; SDS,Shwachman钻石综合征; DC,Dyskeratosis Congenita。仅在专业实验室中进行4个研究测试;在模棱两可或非信息遗传学的情况下有用。5通常在成年人中呈现。6这些IBMF通常表现出多型细胞减少症,并且经常出现其他影响多器官系统的疾病特异性异常。这种区别特征可以帮助将这些条件与DBA综合征区分开,该疾病最初以孤立的红细胞发育不全表现出来。缩写:BM;骨髓; EADA,红细胞腺苷脱氨酶; HBF,胎儿血红蛋白; SLE,全身性红斑狼疮; prca,纯红细胞性植物; CLL,慢性淋巴细胞性白血病; LGL,大颗粒状淋巴细胞性白血病; CT,计算机断层扫描; MRI,磁共振成像; IBMF,继承的骨髓衰竭综合征; FA,Fanconi贫血; SDS,Shwachman钻石综合征; DC,Dyskeratosis Congenita。