XiaoMi-AI文件搜索系统
World File Search System肽聚糖
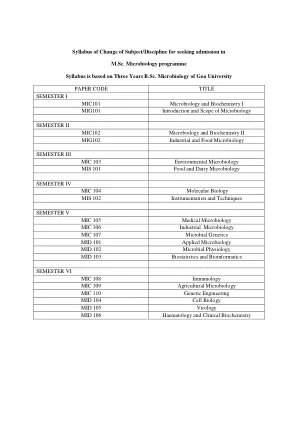
申请入学更改科目/学科的课程大纲
单元 6 大分子 碳水化合物 单糖家族:醛糖和酮糖、三糖、四糖、戊糖和己糖。葡萄糖和果糖的呋喃糖和吡喃糖形式,葡萄糖的 Haworth 投影公式;葡萄糖的椅式和船式。双糖;还原糖和非还原糖的概念,麦芽糖、乳糖和蔗糖的 Haworth 投影。多糖、储存多糖、淀粉和糖原。结构多糖、纤维素、肽聚糖。脂质:储存和结构脂质的定义和主要类别。储存脂质。脂肪酸:结构和功能。必需脂肪酸。三酰甘油结构,结构脂质。磷酸甘油酯:构建块,一般结构。蛋白质:氨基酸,蛋白质的构建块。氨基酸的一般公式和两性离子的概念。蛋白质结构:一级、二级、三级和四级结构。核酸:核苷酸、DNA和RNA的结构;分子生物学中心法则的简要概念。

RIPK2:癌症治疗的一个有希望的靶点
受体相互作用丝氨酸/苏氨酸蛋白激酶2(RIPK2)作为炎症和先天免疫的重要介质,负责将信号传导至细胞内肽聚糖传感器核苷酸寡聚化结构域(NOD)样受体1和2(NOD1/2)下游,从而进一步激活核因子κB(NF- κ B)和丝裂原活化蛋白激酶(MAPK)通路,导致促炎性细胞因子的转录激活并产生炎症反应。因此,NOD2-RIPK2信号通路因其在许多自身免疫疾病中的重要作用而受到广泛关注,使得药物抑制RIPK2成为一种有前途的策略,但对于其在免疫系统之外的作用知之甚少。最近,RIPK2与肿瘤发生和恶性进展有关,迫切需要针对性的治疗。本文主要对RIPK2作为抗肿瘤药物靶点的可行性进行评估,并综述RIPK2抑制剂的研究进展,更重要的是,结合以上内容,分析小分子RIPK2抑制剂应用于抗肿瘤治疗的可能性。

被感染的狗叮咬,伴有动物奈瑟氏菌和奈瑟氏菌,导致隔室综合征,坏死筋膜炎和气候坏死:病例报告
Neisseria是一种革兰氏阴性,催化和氧化酶阳性的球杆菌细菌,能够发酵葡萄糖并产生精氨酸二氢酶[1,2]。是通过释放肽聚糖,脂肪酸糖和外膜外囊泡作为诱导炎症和免疫反应的片段[3]。尽管存在11种已知的奈瑟氏菌种,但人类大多容易受到两种菌株,即脑膜炎和淋病链球菌。neisseria andaryoris和neisseria canis经常与牙龈,口腔和鼻腔分泌犬和猫科动物分泌[2,4]。这些菌株在人类中是罕见的人畜共患病原体,但通常与猫或狗叮咬有关[1]。对这些细菌的非肿瘤人类伤口感染很少见,文献稀疏,病例报告只有14例病例[5]。尚未描述过动物科和猪笼草的病例,引起了隔室综合征和坏死性筋膜炎的坏疽性感染。

MDP
描述Muramyl二肽(MDP)是一种由N-乙酰毛素酸组成的合成免疫反应性肽,其乳酸部分与L-丙氨酸D-异谷胺二肽的N-末端相连。MDP是几乎所有细菌中发现的最小生物活性肽聚糖基序。它首先被识别为Freund完整辅助1中的活动组件。MDP被胞质受体NOD2 2,3识别。配体 - 结合NOD2通过丝氨酸/苏氨酸RIP2(Rick,Cardiak)激酶通过卡片卡均质相互作用4。一旦激活,RIP2介导了NEMO/IKKγ的泛素化,从而导致NF-κB的激活和炎症细胞因子的产生。此外,多泛素化的RIP2募集了TAK1,这会导致IKK复合物激活和MAPK 5的激活。此信号涉及适配器蛋白卡9 6。nod2识别MDP对L-D异构体是立体特异性的,不包括对D-D或L-L类似物3的任何反应。MDP的有效辅助活性也可能与NLRP3炎性症7的激活有关。值得注意的是,已经发现与克罗恩病易感性相关的NOD2突变体在识别MDP 2、3方面缺乏。

周质生物矿化用于半人工光合作用
基于半导体的生物界面通常建立在质膜表面或细胞质内。在革兰氏阴性细菌中,周质空间的特点是封闭且存在大量酶和肽聚糖,为生物矿化提供了额外的机会,从而允许非遗传调节界面。我们通过各种基于电子和 x 射线的成像技术观察到周质内含有单金属和多金属元素的半导体纳米团簇沉淀。周质半导体是亚稳态的,并显示出缺陷主导的荧光特性。出乎意料的是,原位产生的富含缺陷(即低品位)的半导体纳米团簇在与光敏化结合时仍可提高三磷酸腺苷水平和苹果酸的产生。我们扩展了生物混合系统的可持续性水平,包括在初级水平上减少重金属、在次级水平上构建活生物反应器以及在第三级水平上创建半人工光合作用。具有生物矿化功能的周质生物混合体有可能成为各种可持续应用的容错平台。

设计一种基因组减少的细菌来消除体内金黄色葡萄球菌生物膜
细菌为治疗人类疾病提供了一种很有前途的递送系统。在这里,我们设计了基因组减少的人类肺部病原体肺炎支原体作为活生物治疗剂来治疗生物膜相关细菌感染。该菌株具有独特的遗传密码,这会阻碍基因转移到大多数其他细菌属,并且它缺乏细胞壁,这使得它能够表达针对致病菌肽聚糖的蛋白质。我们首先确定去除致病因素可在体内完全减弱底盘菌株。然后,我们设计了合成启动子并确定了内源肽信号序列,当该序列与异源蛋白质融合时,可促进有效分泌。基于此,我们为底盘菌株配备了一个旨在分泌抗生物膜和杀菌酶的遗传平台,从而产生一种能够在体外、离体和体内溶解导管上预先形成的金黄色葡萄球菌生物膜的菌株。据我们所知,这是第一个可以对抗临床相关的生物膜相关细菌感染的工程基因组减少细菌。

幼虫菌群素果蝇成人味反应
动物的生存取决于它们在周围环境中识别威胁的能力。嗅觉,视觉和味觉等感官在取样其生活环境中起着至关重要的作用,包括微生物,其中一些是潜在的致病性。本研究的重点是果蝇味道系统检测细菌的机制。我们证明,形成细菌细胞壁的肽聚糖(PGN)在被成年流量的阵风系统检测时会触发立即进食的厌恶反应。尽管我们将ppk23+和GR66A+脾性神经元视为对PGN的反应的必要性,但我们证明它们在此过程中起着非常不同的作用。时间控制的功能灭活和体内钙成像表明,虽然成年浮球中需要PPK23+神经元直接传播PGN信号,但GR66A+神经元必须在幼虫中起作用才能使未来的成年人变得PGN敏感。此外,当成年流量从轴承条件下饲养的幼虫孵化时,损失了成年浮球对细菌PGN的反应。无菌幼虫的再殖民化,而不是成年人,具有单一细菌的Brevis乳杆菌,是恢复成年人对PGN做出反应能力的足够的能力。我们的数据表明,幼虫的遗传和环境特征对于使未来的成年人有能力应对某些感觉刺激(例如PGN)至关重要。

当主机遇到细胞壁时,反之亦然
肽聚糖(PGN)和相关的表面结构(例如次级聚合物和胶囊)在细菌生理学中具有核心作用。外骨骨骼PGN异聚物是细胞形状的主要决定因素,可使细菌承受细胞质颤音压力。因此,需要高度调节其在细胞生长和分裂过程中的组装,膨胀和重塑,以避免损害细胞存活。同样,组装的调节会影响细菌细胞的形状;不同的形状可以增强不同生态壁ches(例如宿主)中的拟合度。由于细菌细胞壁成分,尤其是PGN,暴露于环境和细菌所特有的环境中,因此在真核生物进化过程中,这些成分已依靠细菌来检测细菌。此外,细胞壁在宿主和微生物之间对话框中的重要信号分子和许多宿主反应的目标中成为重要的信号分子。数百万年的协同进化已导致PGN片段在塑造宿主生理学和建立持久的微生物和宿主之间的持久性共生方面发挥了关键作用。因此,此对话的扰动可能导致诸如慢性炎症疾病之类的病理。同样,病原体制定了复杂的策略来操纵系统以增强其生存和生长。

病原体代谢产物检查点:NHR在后卫上
如何区分有益与微生物的有害相互作用?生物体使用多种方式来了解它们何时被感染。一个人不依赖于直接检测病原体,而是依赖于感染的后果。果蝇有一个惊人的例子,当真菌蛋白酶通过裂解内源性固定前体Spaetzle(2)来激活Toll信号通路(2)。此类检测系统是多种多样的。在秀丽隐杆线虫中,例如,顶端细胞外基质的破坏或核糖体,线粒体或核仁功能的扰动会通过或多或少理解的机制导致防御基因的表达(3,4)。并行,生物具有不同的模式识别受体(PRR),直接识别非自我。例如,在哺乳动物中,这可以通过类似收费的受体(TLR)。这些结合了一系列原型微生物部分,包括革兰氏阴性细菌的LPS。肽聚糖,另一种细菌细胞壁成分,由昆虫和点头样受体中的PGRP受体和哺乳动物中的TLR2检测到。奇怪的是,除了检测病毒双链RNA外,尽管进行了激烈的研究,但在秀丽隐杆线虫中仍缺乏直接病原体识别的例子(3)。Peterson等人的论文。在本期免疫力中,部分填充了该空隙,因为它描述了秀丽隐杆线虫中的新型PRR,秀丽隐杆线虫是一种检测特定有毒细菌代谢物的核激素受体(NHR)。

溶菌酶抑制剂作为溶菌酶分析的工具
摘要:溶菌酶是动物先天免疫系统的通用成分,它们通过水解其主细胞壁聚合物肽聚糖而杀死细菌。已经确定了三个主要的溶菌酶家族,称为鸡(c) - ,鹅(g)和无脊椎动物(i)-type。在反应中,细菌对三个溶菌酶家族中的每一个都进化了特定的蛋白抑制剂。在这项研究中,我们开发了由三个由C-,G-和I型型抑制剂功能化的三个Af-fiential矩阵的序列阵列,用于溶菌酶键入,即检测和区分溶菌酶从动物中流动或提取物。该工具在蓝贻贝(Mytilus Edulis)上进行了验证,其基因组具有多种推定的I-,G-和C型溶菌酶基因。血淋巴等离子体包含I-和G型,但不含C型溶菌酶。此外,分别分析了缺乏或过量产生I-type或G-type溶菌酶抑制剂的嗜水和大肠杆菌菌株的血淋巴存活,以研究两种溶菌酶在先天免疫中的作用。结果表明,G型溶菌酶在蓝贻贝的先天免疫中发挥了积极作用,但未能显示I-type溶菌酶的贡献。使用基于抑制剂的AFINIDE色谱法进行溶菌酶填充将是研究动物先天免疫的有用新工具。