XiaoMi-AI文件搜索系统
World File Search System自然选择

婴儿期,童年和青春期的演变
人类的生活在延长的时钟上发挥作用,从新生儿到繁殖成年人的时间最长(平均十年)在哺乳动物世界中闻名。,即使是蓝鲸或大象,甚至在我们附近的哺乳动物跨越了生命,也只能在五到十年内达到生殖年龄。只有黑猩猩甚至接近我们,在野外十四岁至15岁时就达到了第一胎。“生活缓慢而死亡”可能是我们整体策略的名称,但并不是人类生活的所有特征都适合“缓慢”类别。的新颖性已经发展,包括缩写的婴儿期,拥挤的间隔间隔,在休闲后的依赖后和生长抑郁症时期,我们称为“童年”,以及以新颖的成长反弹为标志的青春期(见Smith and Tompkins 1995)。认识到人类生活课程中的阶段无处不在。全世界的社会,传统和工业,标记生活阶段,并带有通道仪式,可能会引入名称,饮食,外观,居住,关联,特权和责任的变化。出生,童年,青春期,成年和怀孕通常都伴有仪式和文化地位的改变。在文学中,从索伦(Solon)到莎士比亚(Shakespeare)的2,000年,西方作家将人类的生命分为七人类(七年或七个时期),四分之一或十二个,受命理,占星术或宗教文本影响(见Boyd 1980)。我们今天使用的英语术语主要源自拉丁语:女队(无法说话),Adolescere和成人(成长,长大),Iuniores(Young),Senior和Senex(年龄较大,老年)。“孩子”有古老的英语根。到16世纪,它在婴儿期 - 年龄 - 青春期的发展中似乎是自己的阶段。18世纪和十九世纪的学者有时是出于医疗目的,通常通过落叶和恒牙喷发来标出早期的生命阶段(Boyd 1980)。对进化人类学中生活阶段的认识归功于人类成长研究的悠久历史,也归功于生活史的领域,在那里,广泛的“比较方法”是一种核心工具。“生命历史”一词来自自然历史,它指的是物种的生长,发展,规模,生命跨度和生命周期。作为一个研究领域,“生活史”结合了人口统计学,伦理学/民族志学和自然选择理论,以引起一般原则,解释了自然选择在生命周期中的作用,既可以在自然世界中产生各种各样的生命,又可以在新的选择性制度下对人口产生预测。

微生物对不同环境条件的适应
微生物在地球上无处不在,几乎可以栖息在任何环境中。在复杂的异质环境中或面对生态干扰时,微生物通过一系列细胞和分子系统来适应不断变化的环境条件。它们的栖息地各不相同,从南极洲的寒冷微观世界到地热火山区,从陆地到海洋,从高碱性区域到极酸性区域,从淡水到咸水源。多样化的生态微生物生态位归因于微生物在温度、营养物质可用性和 pH 波动下的多功能性、适应性。这些生物已经发展出一系列机制来应对环境变化,从而保持其在调节重要生态系统功能中的作用。在细胞、遗传和分子水平上彻底研究了适应性微生物性质的潜在机制。适应性由一系列过程介导,例如自然选择、基因重组、水平基因转移、DNA 损伤修复和多效性事件。这篇评论论文除了强调不同环境条件下微生物适应的分子网络之外,还提供了有关微生物适应性的基本见解。

进化是事实
进化的概念源自查尔斯·达尔文和阿尔弗雷德·拉塞尔·华莱士的创新思想,他们提出了自然选择理论,解释了物种如何适应和进化。该理论得到了古生物学、生态学、遗传学、发育生物学、分子生物学等多个科学领域的大力支持。证据来自多个方面:从化石记录(显示生命形式随时间的演变)到 DNA(其基因序列揭示了不同生物之间令人印象深刻的相似性,强调了共同的祖先)。例如,将人类 DNA 与黑猩猩的 DNA 进行比较,相似度接近 98.8%,表明它们具有相对较新的共同祖先。此外,该理论还通过物种的地理分布以及适应辐射和适应趋同等过程得到了说明,当祖先群体在不同环境中定居并产生其他物种时就会发生这些过程。比较胚胎学和发育生物学也表明,发育早期的微小基因变异可能导致成年形态和功能的巨大差异。
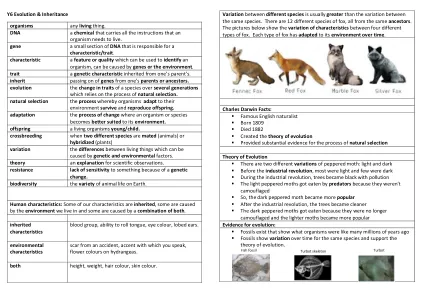
y6进化和继承生物有生物。 DNA ...
生物都有生物。dna一种化学物质,它带有生物体需要生存的所有指示。基因一小部分DNA负责特征/性状。可以用来识别生物体的特征或质量,可能是由基因或环境引起的。特质是从父母的父母继承的遗传特征。继承了父母或祖先的基因传递。进化,几代人在几代人中的特征变化依赖于自然选择的过程。自然选择生物体适应其环境的过程生存并繁殖后代。适应有机体或物种更适合其环境的变化过程。后代生物幼儿/孩子。杂交当两个不同的物种交配(动物)或杂交(植物)变异时,生物之间可能由遗传和环境因素引起的生物之间的差异。理论对科学观察的解释。由于遗传变化而抗性缺乏对某物的敏感性。生物多样性地球上动物生命的多样性。人类特征:我们的某些特征是继承的,有些是由我们生活的环境引起的,有些是由两者的组合引起的。特征示例的类型继承特征

基于引言的增强遗传算法...
抽象的遗传算法是一种灵感的元元素算法,灵感来自自然选择理论,可以解决各种优化问题。本研究提出了一种方法,目的是促进遗传算法的探索和开发。通过调整初始人口并增加一组固定站,可以提高勘探能力。这种修改增加了解决方案人群之间的多样性,这使算法能够从局部最佳距离中逃脱,并在更少的一代中融合到全球最佳最佳。另一方面,为了增强剥削能力,建议增加了所选父母的数量,并提出了相应的跨界技术。在拟议的技术中,在此过程中产生后代的父母数量是可变的,并且可能超过两个。通过检查几个基准功能和工程设计问题,已验证了所提出方法中修改的有效性。关键字:遗传算法;元式优化;固定车站组;可变的多父跨界。收到:2024年1月17日;接受:2024年3月17日

量子启发遗传算法解决组合优化问题
工程、金融和基因组学等众多行业都遇到了组合优化问题。这些问题需要通过从有限的集合中选择最佳组合或排列来优化给定的目标函数,但要受到特定限制(Smith,2010 年)。然而,由于这些问题本身就很复杂,因此有时很难通过计算解决,而且需要很长时间。研究人员已经使用了各种优化策略来解决这些问题,其中遗传算法 (GA) 脱颖而出(Goldberg,1989 年)。在问题的解空间中寻找解决方案是通过遗传算法完成的,遗传算法的灵感来自自然选择和进化的思想。它们使用选择、交叉和突变等遗传运算符在几代中开发出一个潜在解决方案群体(Holland,1975 年)。尽管 GA 能够有效地处理各种优化问题,但 Mitchell(1998 年)发现,在处理具有高维解空间的困难组合优化问题时,它们的性能可能会下降。此外,搜索过程可能会陷入局部最优,这使得找到整体最优解决方案变得更加困难(Vose,1999)。

杂交高粱和自交高粱产量的适应性和可塑性
自交物种中生长时间的延长(Barrett & Charlesworth, 1991)可以解释自交物种中观察到的较低杂种优势水平。杂种优势的程度在物种内测量性状、遗传背景(Tracy & Chandler, 2006)和测试环境(Flint-Garcia et al., 2009; Lippman & Zamir, 2007; Mindaye et al., 2016)之间差异很大。在没有过度遗传漂变或足够基因流的情况下,植物种群倾向于适应来自生物和非生物挑战的人工或自然选择力量,从而导致对环境的局部适应(Janzen et al., 2022; Leimu & Fischer, 2008)。可以在认为当地植物类型相对于外来引进植物具有适应性的环境中测试当地植物和外来植物性能之间的区别(Kawecki & Ebert, 2004)。鉴于遗传分化和杂种优势之间的普遍关联 (Jordan et al., 2003; Moll et al., 1965; Zhang et al., 2010),不同环境中遗传和表型分化的相互作用对于理解和利用多种来源材料的作物育种计划中的杂种优势至关重要。

没有证据表明果蝇中跨代免疫启动
大多数生物都在病原体恒定和反复接触,从而导致持久的自然选择,以实现更有效的抗击感染方法。这可能包括基于内存的免疫的演变,以增加几代人内部和几代人反复遇到的病原体的保护。有不同的证据表明在非魔力脑中缺乏抗体介导的脊椎动物的抗体特征的非魔力。在这项工作中,我们在孕产妇挑战10种不同细菌病原体的果蝇果蝇(Drosophila Melanogaster)的成人后代进行了跨代免疫释放。我们专注于索菲拉(Sophila)的自然机会性病原体,涉及从10%到100%的宿主死亡率范围。我们通过化粪池损伤感染了飞蛾,并测试了成年后代对感染的耐药性增强,以抑制细菌增殖和存活感染的能力。我们将母亲纳入了每个测试的细菌的四个类别:那些幸存的感染,那些屈服于感染的人,无菌伤害的控制和无受伤的控制。我们没有发现任何一类母亲响应任何细菌的跨代启动的证据。

棘鱼个体发育过程中年龄相关的遗传结构
自然选择下性状的可遗传变异是进化反应的先决条件。虽然人们认识到性状的遗传性可能随性状表达的环境条件而随空间和时间的变化,但人们对导致给定性状预期选择反应的遗传变异在个体发育的不同阶段可能变化的可能性知之甚少。具体而言,尚不清楚不同的基因座是否在整个发育过程中影响性状的表达,从而为野外选择提供额外的变异来源。在这里,我们表明,在九刺鱼(Pungitius pungitius)的整个个体发育过程中,体型这一重要的生活史性状都是可遗传的。尽管如此,数量性状基因座分析和跨年龄遗传相关性分析都表明,不同的染色体/基因座在不同的个体发育时间点对这种遗传性有贡献。这表明,体型大小可以在个体发育的不同阶段对选择作出反应,但这种反应是由不同发育点的不同基因座决定的。因此,我们的研究为我们理解个体发育的遗传学提供了重要的结果,并为研究年龄特异性遗传结构作为非平行进化的来源开辟了一条有趣的研究途径。

抽象的。 Aditama R, Tanjung ZA, Sudania WM, Nugroho YA, Utomo C, Liwang T. 2020. 密码子使用偏向性分析揭示了油棕豚鼠的最佳密码子
摘要。Aditama R、Tanjung ZA、Sudania WM、Nugroho YA、Utomo C、Liwang T。2020. 密码子使用偏向分析揭示了油棕中的最佳密码子。生物多样性 21:5331-5337。报道了油棕基因组的密码子使用偏向,采用了几个指标,包括 GC 含量、相对同义密码子使用 (RSCU)、有效密码子数 (ENC) 和密码子适应指数 (CAI)。观察到 GC 含量的单峰分布,并与非草本单子叶植物的特征相匹配。同义密码子使用偏向的对应分析 (COA) 表明主轴由 GC 含量强烈驱动。油棕基因的 ENC 和中性图表明,自然选择在塑造密码子使用偏向方面比突变偏向发挥了更重要的作用。计算出的 CAI 与油棕基因表达的实验数据之间存在正相关性,表明该指数具有良好的能力。最后,十八个密码子被定义为“最佳密码子”,可为异质表达和基因组编辑研究提供有用的参考。