XiaoMi-AI文件搜索系统
World File Search System轴突

BHAC515.PDF- -orca -Cardiff University
在儿童晚期和青春期的白质微观结构发育主要是通过增加轴突密度和髓磷脂厚度来驱动的。ex vivo研究表明,轴突直径的增加会促进人们观察到的青春期发作观察到的轴突密度的增加。在这项横断面研究中,使用超强梯度磁共振成像扫描仪扫描了50名典型的8-18岁参与者。微结构特性,包括明显的轴突直径(D A),髓磷脂含量和G-Ratio,在call体的区域估计。我们观察到D A,髓磷脂含量和G-Ratio的年龄相关差异。在青春期早期,与雌性相比,男性在脾脏中具有更大的d a,并降低了call体的真实和身体的髓磷脂含量。总的来说,这项工作提供了有关发展中轴突直径和发育中的人脑含量的个体差异的发育,青春期和认知相关性的新见解。

DREBRIN调节皮质II/III层中感官神经元中的侧支轴突分支
神经发育障碍(NDDS)是一组复杂的神经系统疾病和精神疾病。功能性和分子成像技术,例如静息状态功能磁共振成像(RS-FMRI)和正电子发射tomog-raphy(PET),可用于在人类和人类模型中成熟期间在成熟期间非侵入性和纵向测量网络活性。Here, we review the current knowledge on rs-fMRI and PET biomarkers in the study of normal and abnormal neurodevelopment, including intellectual disability (ID; with/without epilepsy), autism spectrum disorder (ASD), and attention deficit hyperactivity disorder (ADHD), in humans and rodent models from birth until adulthood, and evaluate the cross-species translational value of the imaging biomarkers.迄今为止,只有少数孤立的研究使用RS-FMRI或PET在婴儿期(神经发育的关键时期)中研究啮齿动物的神经发育(异常)。Further work to explore the feasibility of performing functional imaging studies in infant rodent models is essential, as rs-fMRI and PET imag- ing in transgenic rodent models of NDDs are powerful techniques for studying disease pathogenesis, developing noninvasive pre- clinical imaging biomarkers of neurodevelopmental dysfunction, and evaluating treatment-response in disease-specific models.

巨噬细胞耗尽可恢复DRG微环境并防止硼替佐米诱导的神经病中的轴突变性
。cc-by-nc-nd 4.0国际许可证(未经同行评审证明)获得的是作者/资助者,他授予Biorxiv授予Biorxiv的许可,以永久显示预印本。这是该版本的版权持有人,该版本发布于2025年1月24日。 https://doi.org/10.1101/2025.01.22.634362 doi:Biorxiv Preprint

模块 1——大脑和神经系统简介
一旦神经递质与受体结合,就会发生一系列事件。首先,神经递质携带的信息被传递给接收神经元。其次,神经递质被灭活。它要么被酶分解,要么被释放它的轴突重新吸收。其他分子,称为转运分子,完成这一重新吸收过程。这些分子位于释放神经递质的轴突的细胞膜中。它们从突触中拾取特定的神经递质,并将它们带回细胞膜并进入轴突,在那里它们被回收以供日后使用。请注意,这个过程适用于大多数神经递质,但并非适用于所有神经递质。

成功老龄化和你的大脑
神经细胞或神经元是大脑的主力。它们的纤维或轴突与其他神经元形成突触连接。当神经元被激活时,它会向轴突发送低水平电流,释放大脑化学物质(神经递质),这些化学物质会扩散到微小的间隙中,并附着在接收神经元的受体上。这会引发一系列化学事件,信号沿着轴突传递,就像接力赛中的选手一样。当我们反复表演或体验某件事时,例如练习乐谱,我们会一次又一次地激活相同的突触回路。这些重复提高了回路的效率,并将体验或行为编码为持久记忆。

摘要 PDF 海报
摘要:近端周围神经损伤 (PNI) 需要长距离轴突再生才能实现目标神经支配和运动功能恢复。虽然成熟的周围神经元在受伤后可以缓慢再生受损的轴突,但在慢性失神经支配后,它们往往无法在运动终板上形成功能性突触,导致即使立即进行手术修复也无法完全恢复运动功能。在过去的十年中,人们付出了很多努力来了解受伤后成功轴突再生所需的分子机制。许多再生相关基因 (RAG) 已被确定在轴突再生中起着不可或缺的作用。在这些 RAG 中,已知在受损的视网膜神经节细胞 (RGC) 中同时消融 PTEN 和 SOCS3 可在视神经挤压伤后诱导持续和长距离的轴突再生。尽管基于病毒的基因传递系统近年来作为各种神经退行性疾病的潜在治疗选择得到了迅速发展,但沉默 PTEN 和 SOCS3 等肿瘤抑制基因可能会对致瘤性产生不良影响,从而限制了它们在临床实践中的治疗应用。因此,本研究旨在识别在神经系统损伤后能够诱导强劲轴突再生和功能恢复的生物活性小分子。我们首先从公开的微阵列数据集中识别了 PTEN 和 SOCC3 同时删除的 RGC 中的差异表达基因,并使用该基因表达谱特征查询药物相关基因表达谱数据库 LINCS,以对生物活性小分子进行计算机筛选。使用模式匹配算法,选出 4 种具有高连接得分的生物活性小分子,使用轴突切断的背根神经节 (DRG) 神经元的体外培养进行功能验证。其中,有一种小分子被发现能有效促进体外培养的 DRG 神经元的神经突生长,以及 PNI 小鼠模型中的体内轴突再生。用这种小分子治疗的小鼠在坐骨神经挤压伤后感觉和运动功能均得到了早期恢复。这些小鼠的复合肌肉动作电位 (CMAP) 幅度也显著增大
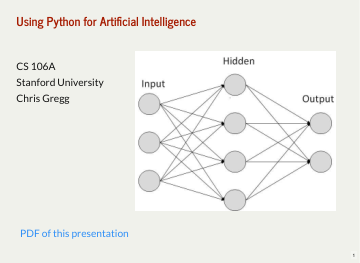
使用 Python 实现人工智能
生物神经元有三种主要组成部分:树突、胞体(或细胞体)和轴突。树突接收来自其他神经元的信号。胞体汇总传入的信号。当接收到足够的输入时,细胞就会激发;也就是说,它会通过轴突向其他细胞传输信号。

701587 HT DNA 5K标记701637 HT DN
图2:Netri神经流体设备中神经毒性扩散测定的开发。主要大鼠神经元在双链偏移Neobento™中生长,以允许细胞和突触的分隔,并用MAP2和DAPI标记。使用10X目标以共聚焦模式在Operetta CLS系统上获取图像。a)基于轴突腔中的βO浓度的增加,ETAP-LAB能够证明体细胞腔中的核数量减少以及轴突腔中轴突的碎片(概述图像)。b)车辆控制中健康神经元的详细视图。

在22q11.2缺失综合征
22Q11.2缺失综合征或22q11.2ds是一种遗传综合征,除了整个大脑的广泛结构和功能异常外,还与高精神分裂症和自闭症谱系障碍相关。实验动物模型已鉴定出神经元连通性的降低,例如,轴突分支的轴突长度和复杂性降低,是22q11.2ds中非典型大脑发育的主要机制。然而,尚不清楚在22q11.2ds的人中是否还可以观察到轴突形态中的差异。在这里,我们在22q11.2ds(12 - 15年)的参与者中提供了无与伦比的在体内表征,并使用对轴突形态敏感的定制磁共振成像扫描仪进行了典型发育(8-18岁)。提取了丰富的扩散MRI指标,以呈现典型和非典型白质发育的微结构pro插曲,并提供了22q11.2ds个人连通性差异的新证据。最新的大规模财团研究22q11.2ds鉴定出较高的扩散各向异性,并降低了水的总体扩散迁移率,这是个体跨年龄范围内白质的标志性微结构变化(6 - 52岁)。我们观察到本研究中包括的白质区域中的类似发现,除了确定轴突形态学中的缺失。这与减少的道量测量值结合了以下假设:22q11.2ds中异常的微观结构连通性可能是通过密集的轴突介导的,其直径不成比例。我们的发现提供了对22q11.2ds体内白质表型的见解,并促进对神经发育和精神疾病中共同特征的持续调查。

研究衰老schwann细胞的神经修复能力的研究进展
周围神经损伤(PNI)与老年患者受伤神经的延迟修复有关,导致神经功能,慢性疼痛,肌肉萎缩和永久残疾的丧失。因此,应研究衰老患者周围神经延迟修复的基础机制。schwann细胞(SCS)在修复PNI和调节损伤后各种神经治疗基因方面起着至关重要的作用。sc还通过各种方式促进周围神经修复,包括介导神经脱髓鞘,分泌神经营养因素,建立büngner带,清除轴突和髓磷脂碎屑以及促进轴突雷格尔。然而,年龄的SC经历了结构和功能变化,导致脱髓鞘和去分化障碍,神经营养因子的分泌减少,轴突和髓磷脂碎屑的清除受损以及轴突再髓鞘的能力降低。结果,老化的SC可能会导致受伤后神经修复的延迟。本评论文章旨在研究衰老SC的神经修复能力降低的机制。