XiaoMi-AI文件搜索系统
World File Search System重排
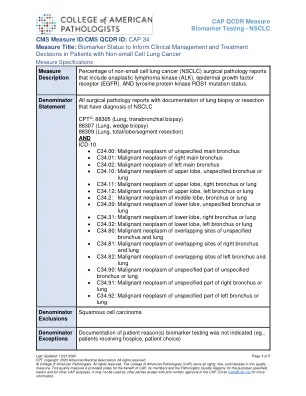
CAP 34 措施标题:生物标志物状态用于指导临床...
这一重要信息对于协调护理和有效利用资源至关重要。因此,了解 ROS1、ALK 和 EGFR 的突变状态对于晚期 NSCLC 的适当临床决策是必不可少的。如果在一线化疗之前或期间发现一种或多种这些重排,则应考虑替代疗法。1% 至 2% 的非小细胞肺癌会出现 ROS1 重排,并且可预测对一线治疗药物克唑替尼和色瑞替尼疗法的反应。反应率(包括完全反应)接近 70% (2-4)。对于 ALK1,除了识别可能对靶向疗法有反应的肿瘤外,了解 ALK 重排状态通常可预测对免疫疗法的反应较差。大约 5% 的肺腺癌具有涉及 ALK 基因的染色体重排,并与 ALK 蛋白过表达有关。患有此类肿瘤的患者对 ALK 酪氨酸激酶抑制剂(如克唑替尼)的治疗有反应 (2, 5)。最后,约 20% 的肺腺癌含有 EGFR 激活突变,这预示着对 EGFR 酪氨酸激酶抑制剂(如厄洛替尼)的治疗有反应 (2, 6-10)。

晚期非小细胞肺癌 (NSCLC) 中的 NTRK 和 NRG1 基因融合
随着个性化医疗的兴起,非小细胞肺癌也开始出现。自 2004 年发现表皮生长因子受体 (EGFR) 突变以来,从分子水平上定义的可从靶向治疗中获益的患者亚组名单已大幅增加,目前的国际指南建议对所有新诊断的局部晚期或转移性非鳞状非小细胞肺癌患者进行至少 5-8 种生物标志物的分子检测,以选择最佳患者 (5-9)。非小细胞肺癌中 ALK 重排的惊人故事促使人们寻找其他可能用于靶向治疗的致癌重排。神经调节蛋白 - 1 (NRG1) 和 NTRK 融合是最近在非小细胞肺癌中发现的两种重排,是肿瘤不可知生物标志物的两个杰出例子。虽然这两种基因异常相对罕见,但它们代表了 NSCLC 的两个临床相关亚组,可以从靶向治疗中获益。本文我们全面概述了 NRG1 和 NTRK 重排 NSCLC 的生物学和临床病理学特征,以及关于利用这些靶点进行治疗的现有数据。
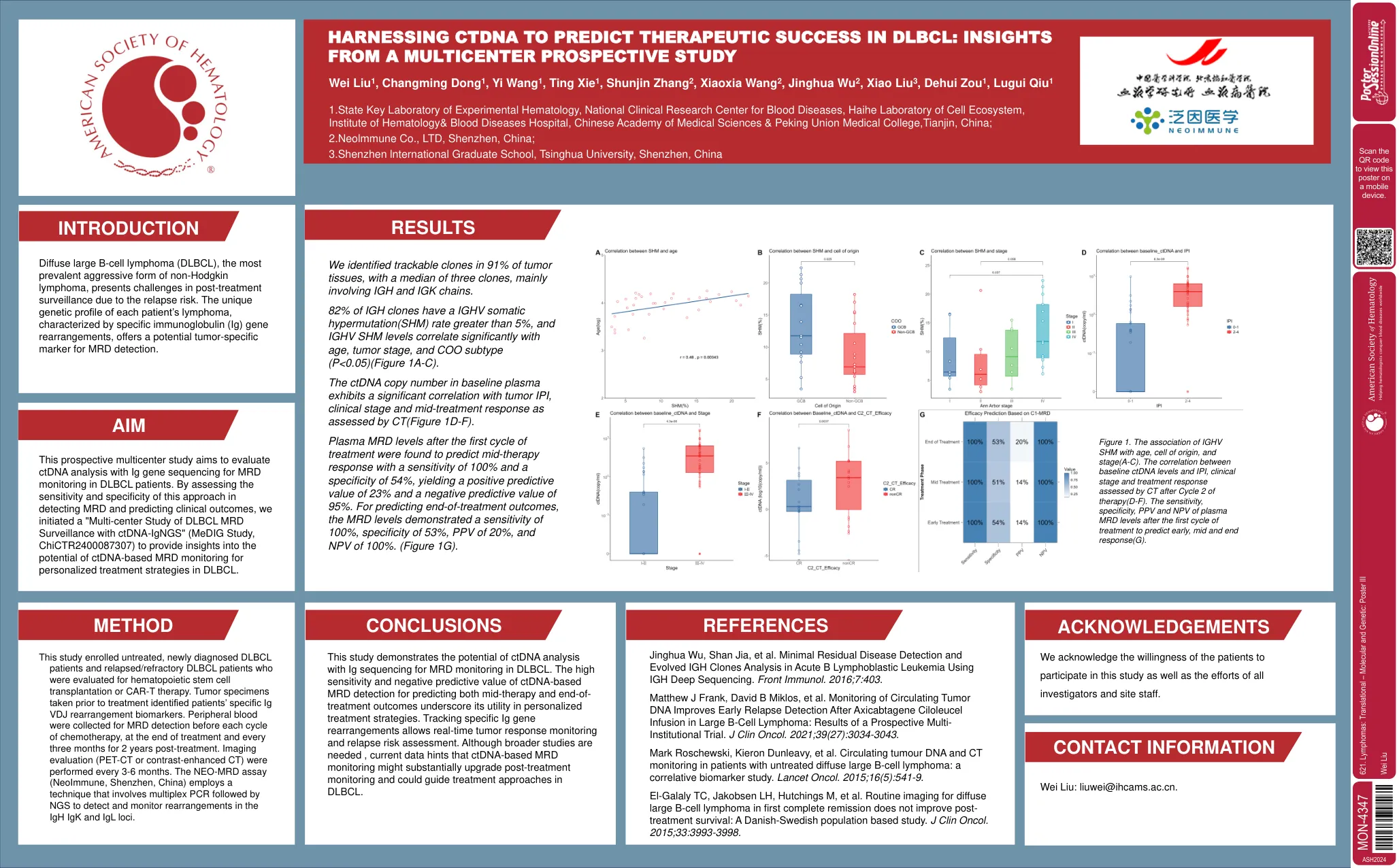

诊所心力衰竭的干细胞疗法
尽管在理解肿瘤的病因和生物学以及增强免疫学控制和引入晚期治疗方式的作用方面取得了重大进展,但肺癌仍然是全球癌症相关死亡的主要原因(1,2)。 非小细胞肺癌(NSCLC)是最常见的肺癌类型,占所有肺癌诊断的84%(3,4)。 尽管化疗仍然是一种常见的治疗方法(3,5),但可靶向的驱动基因突变彻底改变了晚期NSCLC患者的治疗景观。 转染期间重新排列(RET)原始癌基因编码参与正常胚胎发育的跨膜受体酪氨酸激酶。 ret基因融合或重排发生在1% - 2%的NSCLC病例(6)中,尤其是在年轻人(≤60岁)中,不吸烟患有腺癌患者。 这些重排可能会使癌症对某些化学疗法药物的反应更敏感,例如Pemetrexed(7)。 肺腺癌中的恶性胸膜积液(MPE)经常重排(8)。 建议检测RET改变,以识别可能有资格获得RET抑制剂的NSCLC患者。 可用于检测RET重排的分子测试技术包括下一代测序(NGS),逆转录聚合酶链反应(RT-PCR),原位杂交(FISH)和免疫组织化学(9)以及分析循环肿瘤DNA(CTDNA)(CTDNA)(CTDNA)(10)(10)(10)。 selpercatinib是一种新型的酪氨酸激酶抑制剂,它是RET蛋白及其变体活性的选择性阻滞剂。肺癌仍然是全球癌症相关死亡的主要原因(1,2)。非小细胞肺癌(NSCLC)是最常见的肺癌类型,占所有肺癌诊断的84%(3,4)。尽管化疗仍然是一种常见的治疗方法(3,5),但可靶向的驱动基因突变彻底改变了晚期NSCLC患者的治疗景观。转染期间重新排列(RET)原始癌基因编码参与正常胚胎发育的跨膜受体酪氨酸激酶。ret基因融合或重排发生在1% - 2%的NSCLC病例(6)中,尤其是在年轻人(≤60岁)中,不吸烟患有腺癌患者。这些重排可能会使癌症对某些化学疗法药物的反应更敏感,例如Pemetrexed(7)。肺腺癌中的恶性胸膜积液(MPE)经常重排(8)。建议检测RET改变,以识别可能有资格获得RET抑制剂的NSCLC患者。可用于检测RET重排的分子测试技术包括下一代测序(NGS),逆转录聚合酶链反应(RT-PCR),原位杂交(FISH)和免疫组织化学(9)以及分析循环肿瘤DNA(CTDNA)(CTDNA)(CTDNA)(10)(10)(10)。selpercatinib是一种新型的酪氨酸激酶抑制剂,它是RET蛋白及其变体活性的选择性阻滞剂。通过抑制RET,selpercatinib有助于破坏促进存在RET改变的细胞中癌症生长和存活的信号传导途径(11)。我们报告了一个具有挑战性的病例,即在接受化学免疫疗法治疗失败后,在第二行治疗的患者在第二行治疗。

MRIADMYCHOICE®CDX加上技术规格...
•用于体外诊断•仅用于专业用途•仅用于处方使用•MyChoice®CDXPlus测定法可以识别肿瘤中的种系和躯体变体,但没有区分两者。•等位基因频率的变化低于已建立的检测极限。•跨越长插入和缺失(Indels)或重排的DNA片段的杂交效率降低可能导致最终测序文库中突变DNA分子的代表性不足。这将导致观察到的序列读取频率跨越突变的频率。•Indels> 25 bp的长度可以通过MyChoice®CDXPlus分析检测到。但是,检测到任何特定indel的能力可能会受到突变的位置和性质,局部序列上下文,DNA质量以及所提供的样品中非肿瘤DNA含量的影响。•MyChoice®CDXPlus分析已设计为检测基因组重排,包括涉及启动子和分析基因的编码外显子的大重排(LRS);但是,

使用 CRISPR/Cas9- 改组酵母基因组...
尽管转座因子之间的同源重组可通过促进染色体重排来驱动酵母基因组进化,但其潜在机制的细节尚未完全阐明。在酿酒酵母基因组中,最常见的转座子类别是逆转录转座子 Ty1。在本文中,我们探讨了 Cas9 诱导的针对 Ty1 因子的双链断裂 (DSB) 如何在该酵母物种中产生基因组改变。在 Cas9 诱导后,我们观察到染色体重排(例如缺失、重复和易位)显著增多。此外,我们发现有丝分裂重组率升高,导致杂合性丧失。通过 Southern 分析结合短读和长读 DNA 测序,我们揭示了逆转录转座子中诱导的重组的重要特征。几乎所有的染色体重排都反映了 Ty1 元件处 DSB 的修复,这是通过非等位基因同源重组实现的;成簇的 Ty 元件是染色体重排的热点。相反,大部分(约四分之三)等位基因有丝分裂重组事件在独特序列中存在断点。我们的分析表明,后一些事件反映了 Ty 元件中产生的断端的广泛处理,这些断端延伸到独特序列中,从而导致断裂诱导的复制。最后,我们发现单倍体和二倍体菌株对用于修复双链 DNA 断裂的途径有不同的偏好。我们的研究结果表明,逆转录转座子中的 DNA 损伤在推动基因组进化方面的重要性。

来自2,7-二溴-9,9'-Bianthryl的蒽融合的锯齿烯纳米纤维的地下合成揭示了意外的环重排
摘要:地下合成已成为一种有力的策略,用于制造原子上精确的石墨烯纳米骨(GNR)的前所未有的形式。但是,锯齿形GNR(ZGNR)的地下合成仅取得了有限的成功。在此,我们报告了2,7-二溴-9,9' - 苯甲酰基的合成和表面反应,作为朝向π-延伸ZGNRS的前体。通过扫描隧道显微镜和高分辨率非接触原子力显微镜的表征清楚地证明了烟碱融合的ZGNR的形成。独特的骨骼重排,可以通过分子内多尔 - alder cycloadition来解释。对蒽接受ZGNR的电子特性的理论计算显示自旋极晶状体和0.20 eV的狭窄带隙。关键字:地下合成,石墨烯纳米替恩,表面反应,重排,边缘状态■简介

EVI1控制KDM6B介导的组蛋白去甲基化为...
等离子体,单核细胞,中性粒细胞或血小板的增殖增加(1、3、4)。大约30%的被诊断为MD的患者最终患有急性髓样白血病(AML)(5)。eVI1首先被鉴定为具有逆转录病毒诱导的髓样恶质的小鼠中生态病毒整合的常见位点(6)。人类EVI1(MECOM)基因位于Chro-Mosome 3Q26上,EVI1的多种同工型在MECOM基因座(7)中编码。3q26染色体的重排,导致EVI1的上调,经常发生在包括MDS,AML和慢性髓样白血病(CML)在内的髓样恶性疾病中(8-10)。MDS,AML和CML具有INV(3)/T(3; 3)重排通常具有相似的病理特征,预后不良(8、11、12)。It was reported that chromosome rear- rangements cause overexpression of EVI1 due to relocation of enhancers, including GATA binding protein 2 (GATA2) enhancer in inv(3)/t(3;3) (q21q26) (13, 14) and MYC super-enhancer in t(3;8) (q26;q24) close to the EVI1 gene (15).EVI1过表达可能发生在没有3染色体重排的MDS患者中。EVI1上调


重离子诱导缺失与拟南芥必需基因分布相关的基因组学研究
重离子束是一种电离辐射,它已作为一种强诱变剂应用于植物育种,并且是一种诱导大规模缺失和染色体重排的有前途的工具。重离子辐照的有效性可以用线性能量转移 (LET;keV µm -1 ) 来解释。不同 LET 值的重离子束会诱发不同类型和大小的突变。已有研究表明,缺失大小随 LET 值的增加而增大,较高的 LET 辐射会诱发复杂的染色体重排。在本研究中,我们将在拟南芥突变体中检测到的重离子束诱导的缺失定位到其基因组中。我们发现,不同的 LET(100 至 290 keV mm -1 )之间的缺失大小相似,其上限受必需基因分布的影响,并且检测到的染色体重排避免了破坏必需基因。我们还重点研究了串联基因 (TAG),即基因组中两个或多个同源基因相邻。我们的结果表明,100 keV µm -1 的 LET 足以破坏 TAG,并且必需基因的分布会强烈影响与其重叠的突变的遗传性。我们的研究结果提供了拟南芥基因组中大量缺失诱导的基因组视图。