XiaoMi-AI文件搜索系统
World File Search SystemATPase
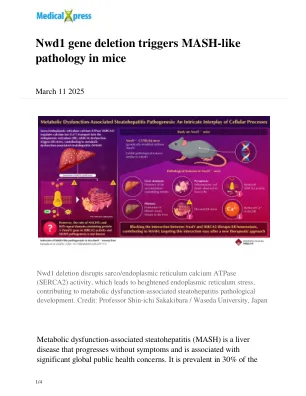
NWD1基因缺失触发小鼠中的含Mash样病理
ER中展开或错误折叠的蛋白的积累激活了一系列称为ER应力的稳态反应,并且持续的ER应力在MASH的发展中起作用。最近的研究表明,调节Ca 2+向ER的Ca 2+转运的Sarco/ER钙ATPase(SERCA2)的功能障碍也可能引起ER应力,从而导致MASH。

2022关于威尔逊病的实践指南
缩写:AASLD,美国肝病研究协会; AI,足够的摄入量; AIH,自身免疫性肝炎; ALF,急性肝衰竭;阿里,急性肝损伤; Alt,丙氨酸氨基转移酶; AST,天冬氨酸氨基转移酶; ATP7B,ATPase Copper运输Beta(基因); ATPase,腺苷三磷酸酶; CCA,胆管癌; CDG,糖基化先天性疾病;中枢神经系统,中枢神经系统; CUEXC,可交换铜; ECT,电抽搐治疗; FDA,食品和药物管理局; HCC,肝细胞癌; INR,国际标准化比率; KF,Kayser - Fleischer; MDD,主要抑郁症; MDR3,多药抗性蛋白3; Mednik,智力(精神)残疾,肠病,耳聋,神经病,鱼质病,角膜疾病;梅尔德,末期肝病的模型; MRI,磁共振成像; NCC,非谷质蛋白 - 结合铜; NWI,新威尔逊指数; PGM1,磷酸葡萄糖酶1; RD,注册营养师; RDA,建议饮食津贴; TTM,四酚糖; UWDR,统一的威尔逊疾病评级量表; WD,威尔逊病。


HEK-293细胞系中的定量分析
不可消除的细胞会产生胞质Ca 2+信号,以响应G蛋白偶联受体和生长因子受体的刺激(Berridge等,2003; Clapham,2007)。通常,在没有外部Ca 2+的情况下,可以在短时间内观察到这些Ca 2+信号,这表明胞质Ca 2+浓度([[Ca 2+] Cyt)的潜在增加的主要机制是Ca 2+从内质含量网状(Barak和Parak and Parekh,Parekh,2020)中释放出Ca 2+的释放。Along with mitochondria, the clearance of cytosolic Ca 2+ by the plasma membrane Ca 2+ ATPase (PMCA) and the Na + -Ca 2+ -exchanger (NCX) reduces the amount of Ca 2+ that is available to the sarco/endoplasmic reticulum Ca 2+ ATPase (SERCA) for re fi lling ER Ca 2+ stores after each Ca 2+ spike, and as a结果,Ca 2+信号在无Ca 2+的解决方案中(Barak and Parekh,2020年)后降低。为了产生稳定的高胞质Ca 2+尖峰,因此是必不可少的,细胞外Ca 2+的大孔是必须的,这是由商店经营的Ca 2+进入(SOCE)实现的,之所以称为刺激触发的刺激触发,从而降低ER Ca 2+水平(Putney,2017; Lewis,2017; Lewis,2020 2020)。通常,SOCE生成

使用双重 BRG1/BRM(SMARCA4/2)抑制剂 FHD-286 治疗可消除前列腺癌中的肿瘤相关雄激素反应元件 (ARE)
雄激素受体 (AR) 顺反组在前列腺细胞身份的形成中至关重要,而其失调会促进前列腺癌的发展。先驱转录因子 Forkhead box A1 (FOXA1) 已被证明对 AR 募集到雄激素反应元件 (ARE) 至关重要,从而允许对 AR 顺反组进行重新编程,导致前列腺细胞转化。FHD-286 是一种 BRM/BRG1 双 ATPase 抑制剂,目前正在进行 AML 临床试验。在这里,我们表明用 FHD-286 治疗前列腺癌细胞系会导致肿瘤相关 AR 结合位点 (T-ARBS) 处 FOXA1 介导的 ARE 消融。双 ATPase 治疗随后降低了各种致癌 AR 靶基因的表达水平,导致肿瘤细胞活力下降。患者来源的类器官和体内研究均通过显示肿瘤生长减少提供了进一步的验证。令人惊讶的是,抑制 BAF 复合物活性可绕过去势和恩杂鲁胺治疗后常见的 AR 抗性机制,因为含有 AR-V7 剪接变体和神经内分泌类器官的细胞系表现出敏感性。总之,我们的数据说明了通过使用 FHD-286 治疗抑制肿瘤相关 ARE 来治疗 AR 介导的前列腺癌的一种新机制。

pH指纹测定法以鉴定多种经过验证和潜在抗疟药靶标的抑制剂
摘要:需要采用新型作用方式的新药来保护疟疾治疗。近年来,已经测试了数百万种化合物的能力,以抑制无性血液阶段恶性疟原虫寄生虫的生长,从而鉴定出数千种具有抗血流活性的化合物。确定抗白潮化合物的作用机制可以为它们的进一步发展提供了依据,但仍然具有挑战性。相对较高的化合物比例被确定为杀死无性血液阶段的寄生虫,这表明靶向寄生虫的质膜Na + -truding,H + - 进口泵,PFATP4。PFATP4的抑制剂会导致寄生虫的内部[Na +]和pH的特征变化。 在这里,我们设计了一种“ pH指纹”测定法,该测定法可以鲁side pFATP4抑制剂,同时允许检测乳酸抑制剂(和区分)乳酸抑制剂:H +转运蛋白PFFNT,PFFNT,这是验证的抗微药物靶标,以及V型h + h + aTPase的cansy 7,这是一个可能的cansy and and and and typ as and and and and sys。 在我们的pH指纹测定和随后的次要测定中,ZY19489没有显示出V型H + ATPase抑制pH调节的证据,这表明它在寄生虫中具有不同的作用方式。 pH指纹测定也有潜力鉴定蛋白团,酸加载Cl-转运蛋白(S)的抑制剂(对于分子识别(IES)仍然流行),并通过抑制葡萄糖转运蛋白转运蛋白PFHT PFHT或糖溶解作用而起作用。PFATP4的抑制剂会导致寄生虫的内部[Na +]和pH的特征变化。在这里,我们设计了一种“ pH指纹”测定法,该测定法可以鲁side pFATP4抑制剂,同时允许检测乳酸抑制剂(和区分)乳酸抑制剂:H +转运蛋白PFFNT,PFFNT,这是验证的抗微药物靶标,以及V型h + h + aTPase的cansy 7,这是一个可能的cansy and and and and typ as and and and and sys。在我们的pH指纹测定和随后的次要测定中,ZY19489没有显示出V型H + ATPase抑制pH调节的证据,这表明它在寄生虫中具有不同的作用方式。pH指纹测定也有潜力鉴定蛋白团,酸加载Cl-转运蛋白(S)的抑制剂(对于分子识别(IES)仍然流行),并通过抑制葡萄糖转运蛋白转运蛋白PFHT PFHT或糖溶解作用而起作用。因此,pH指纹测定提供了一个有效的起点,使一定比例的抗白质化合物与其作用机理相匹配。关键词:疟疾,恶性疟原虫,药物靶标,pH调节,离子稳态

临床特征和诱导多能干细胞(IPSC)疾病模型的Harel-Yoon综合征模型由复合杂合ATAD3A变体引起
ATPase家族AAA含有域的蛋白3a(ATAD3A)富含线粒体膜,对于维持线粒体结构和功能至关重要。 ATAD3A基因的变体可以导致Harel Yoon综合征(Hayos),这是神经,心血管和其他系统的发育缺陷。 这项研究旨在从患者的体细胞(Zjuchyli001-A)和阴性对照(Zjuchyli002-A)中开发出诱导的多能干细胞(IPSC),作为对ATAD3A变异疾病的病因的进一步研究的有效工具。 我们描述并分析了Proband及其家人的临床表现。 从概率和阴性对照中收集并将其重新编程为IPSC。 此外,我们测量了IPSC中的ATAD3A表达水平,以确认这些细胞系的有效性。 Proband和她的姐姐都病重病,并有复合杂合的ATAD3A变体(F459S/T498NF*13)。 他们的父母是这些变体的载体,没有任何临床表现。 两个变体都位于ATAD3A蛋白的ATPase结构域上。 细胞系Zjuchyli001-A和Zjuchyli002-A呈现多能干细胞的典型特征。 与Zjuchyli002-A相比,Zjuchyli001-A的ATAD3A表达水平显着降低。 这项研究从ATAD3A的复合杂合变体的患者中产生了IPSC,并且是负面对照,作为阐明ATAD3A变体相关疾病的分子机制的有价值工具。ATPase家族AAA含有域的蛋白3a(ATAD3A)富含线粒体膜,对于维持线粒体结构和功能至关重要。ATAD3A基因的变体可以导致Harel Yoon综合征(Hayos),这是神经,心血管和其他系统的发育缺陷。 这项研究旨在从患者的体细胞(Zjuchyli001-A)和阴性对照(Zjuchyli002-A)中开发出诱导的多能干细胞(IPSC),作为对ATAD3A变异疾病的病因的进一步研究的有效工具。 我们描述并分析了Proband及其家人的临床表现。 从概率和阴性对照中收集并将其重新编程为IPSC。 此外,我们测量了IPSC中的ATAD3A表达水平,以确认这些细胞系的有效性。 Proband和她的姐姐都病重病,并有复合杂合的ATAD3A变体(F459S/T498NF*13)。 他们的父母是这些变体的载体,没有任何临床表现。 两个变体都位于ATAD3A蛋白的ATPase结构域上。 细胞系Zjuchyli001-A和Zjuchyli002-A呈现多能干细胞的典型特征。 与Zjuchyli002-A相比,Zjuchyli001-A的ATAD3A表达水平显着降低。 这项研究从ATAD3A的复合杂合变体的患者中产生了IPSC,并且是负面对照,作为阐明ATAD3A变体相关疾病的分子机制的有价值工具。ATAD3A基因的变体可以导致Harel Yoon综合征(Hayos),这是神经,心血管和其他系统的发育缺陷。这项研究旨在从患者的体细胞(Zjuchyli001-A)和阴性对照(Zjuchyli002-A)中开发出诱导的多能干细胞(IPSC),作为对ATAD3A变异疾病的病因的进一步研究的有效工具。我们描述并分析了Proband及其家人的临床表现。从概率和阴性对照中收集并将其重新编程为IPSC。此外,我们测量了IPSC中的ATAD3A表达水平,以确认这些细胞系的有效性。Proband和她的姐姐都病重病,并有复合杂合的ATAD3A变体(F459S/T498NF*13)。他们的父母是这些变体的载体,没有任何临床表现。两个变体都位于ATAD3A蛋白的ATPase结构域上。细胞系Zjuchyli001-A和Zjuchyli002-A呈现多能干细胞的典型特征。与Zjuchyli002-A相比,Zjuchyli001-A的ATAD3A表达水平显着降低。这项研究从ATAD3A的复合杂合变体的患者中产生了IPSC,并且是负面对照,作为阐明ATAD3A变体相关疾病的分子机制的有价值工具。

粘合素超元在循环挤出过程中DNA
抽象的粘着蛋白将基因组DNA挤压成促进染色质组装,基因调节和重组的环。在这里,我们表明粘着蛋白将负超胶引入挤出的DNA中。超螺旋需要粘蛋白的ATPase头,这些头部夹紧DNA以及在粘蛋白的铰链上的DNA结合位点,表明在铰链和夹具之间约束粘蛋白超侧Coil DNA。我们的结果表明,一旦粘蛋白在超涂层期间达到其失速扭矩,DNA挤出会停止,而粘蛋白突变体预测会停滞在较低的扭矩形成细胞中的较短环。这些结果表明,超涂层是环挤出机制的组成部分,并且粘着蛋白不仅通过循环DNA,而且通过将其超级旋转来控制基因组结构。真核间相细胞中的主要文本,SMC(“染色体的结构维持”)复合粘着蛋白将基因组DNA折叠成环和拓扑结构域(TADS;参考(1-4)),可以调节转录(5),重组(6,7),姐妹染色单体分离(8)和复制(9)。粘着蛋白通过由ATP结合 - 水溶液周期控制的构象变化(12)(在(13)中进行了综述),将DNA挤压为环(10,11)。这些是由粘蛋白的SMC1和SMC3亚基催化的,其中包含50 nm长的盘绕螺旋,二聚体“铰链”结构域和球形ATPase'heads'(图s1a),与ABC转运蛋白相关(14)。在ATP结合后,粘蛋白的头部接合和一个称为NIPBL“夹具” DNA的亚基在接合的ATPase头顶上(参考(12,15-17);如图。s1b)。这些动作产生〜15 pn力(18)和循环挤出步骤〜40 nm(100-200 bp;ref。(19)),表明在头部互动过程中将DNA卷入形成循环中。相比之下,在环挤出过程中DNA的构象变化知之甚少。拓扑异构酶II在粘着蛋白环的底部结合并切割DNA(20-23),这表明DNA在这些位点上是超螺旋的。有丝分裂SMC复合物冷凝蛋白还与拓扑异构酶(24-30)共定位并相互作用,并且可以在体外超涂DNA(31-33)。已经提出了此过程发生在循环挤出过程中(31,33),但发现粘着蛋白不适合
FHD-286,BRG1/BRM的有效抑制剂(... ) 发现选择性BRM(SMARCA2)ATPase抑制剂... 靶向蛋白质降解和染色质调节系统 选择性CBP和EP300 DEGRADERS的发现和表征
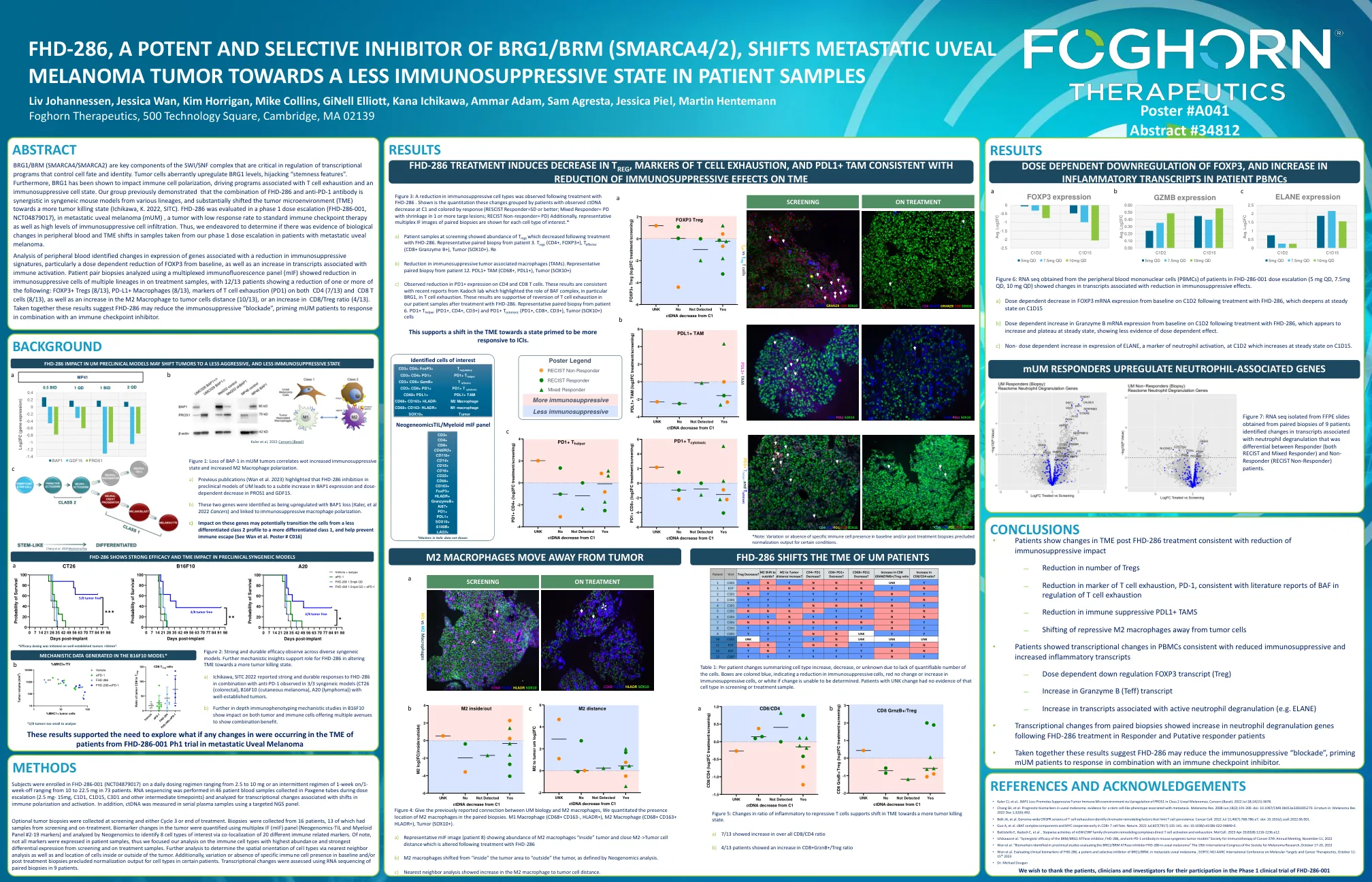
BRG1/BRM(SMARCA4/SMARCA2)是SWI/SNF复合物的关键组成部分,在调节控制细胞命运和身份的转录程序中至关重要。肿瘤细胞异常上调BRG1水平,劫持了“茎特征”。此外,BRG1已被证明会影响免疫细胞极化,驱动与T细胞耗尽和免疫抑制细胞状态相关的程序。我们的小组先前证明,FHD-286和抗PD-1抗体的组合在来自各个谱系的合成小鼠模型中是协同的,并且将肿瘤微环境(TME)实质上转移到了更肿瘤的杀伤状态(Ichikawa,K. 2022,SITC)。FHD-286在1期剂量升级(FHD-286-001,NCT04879017)中评估,转移性卵生黑色素瘤(MUM)是一种对标准免疫检查点疗法的缓解率较低的肿瘤(MUM),以及高水平的免疫抑制细胞抑制。因此,我们努力确定是否有证据表明在转移性紫veal瘤患者中,从我们的1期剂量升级中取出的样品中外周血和TME转移的生物学变化。

表Si.定量RT-PCR的序列序列。
a,siRNA名称/方向序列atp6v1b1_#1 sense 5' - gacaacuucgccaucgucu-3'反义5'-agacgauggcgaaguugu -3'atp6v1b1_#2 ′ B, qPCR primers Gene/direction Sequence ATP6V1B1 Fw 5 ′ -CAGCAGGCTCAGACACTGG-3 ′ Rev 5 ′ -CCCAGGCCTGCTGTCTATCTC-3 ′ Cyclin D1 Fw 5 ′ -CCGTCCATGCGGAAGATC-3 ′ Rev 5 ′ -ATGGCCAGCGGGAAGAC-3 ′ p21 Fw 5 ′ -AGTCAGTTCCTTGTGGAGCC-3 ′ Rev 5 ′ -CATTAGCGCATCACAGTCGC-3 ′ GAPDH Fw 5 ′ -AGAAGGCTGGGGCTCATTTG-3 ′ Rev 5 ′ -AGGGGCCATCCACAGTCTTC-3 ′ AccuTarget Negative Control siRNAs, catalogue no.SN-1013(Bioneer Corporation)。fw,前进; REV,反向; siRNA,小干扰RNA; ATP6V1B1,ATPase H+运输V1亚基B1。