XiaoMi-AI文件搜索系统
World File Search SystemCoV

南亚疫苗接种的COVID状况
摘要背景:冠状病毒(COV)是一种新兴的人类病原体,在世界范围内引起严重的急性呼吸道综合征(SARS)。早期对SAR的生物标志物的鉴定可以促进检测并降低疾病的死亡率。 因此,通过集成的网络分析和结构建模方法,我们旨在探索冠状病毒药物SARS的潜在药物靶标和候选药物。 方法:使用Limma进行了COV感染宿主基因(HGS)表达谱的差异表达(DE)分析。 高度整合的De-cov-HG被选择构建蛋白质蛋白质相互作用(PPI)网络。 结果:使用WalkTrap算法高度互连模块包括模块1(202个节点);从PPI网络检索模块2(126个节点)和模块3(121个节点)模块。 MYC,HDAC9,NCOA3,CEBPB,VEGFA,BCL3,SMAD3,SMAD3,SMURF1,KLHL12,CBL,ERBB4和CRKL被鉴定为潜在的药物靶标(PDT),这些药物靶标(PDT)在COV感染后在人类呼吸系统中高度表达。 功能项生长因子受体结合,C型凝集素受体信号传导,白介素1介导的信号传导,通过MHC I类的TAP依赖性抗原处理以及肽抗原的呈现,刺激T细胞受体信号传导,刺激性T细胞受体信号传导以及先天的响应信号通路,信号传输和细胞座的免疫通道富集。早期对SAR的生物标志物的鉴定可以促进检测并降低疾病的死亡率。因此,通过集成的网络分析和结构建模方法,我们旨在探索冠状病毒药物SARS的潜在药物靶标和候选药物。方法:使用Limma进行了COV感染宿主基因(HGS)表达谱的差异表达(DE)分析。高度整合的De-cov-HG被选择构建蛋白质蛋白质相互作用(PPI)网络。 结果:使用WalkTrap算法高度互连模块包括模块1(202个节点);从PPI网络检索模块2(126个节点)和模块3(121个节点)模块。 MYC,HDAC9,NCOA3,CEBPB,VEGFA,BCL3,SMAD3,SMAD3,SMURF1,KLHL12,CBL,ERBB4和CRKL被鉴定为潜在的药物靶标(PDT),这些药物靶标(PDT)在COV感染后在人类呼吸系统中高度表达。 功能项生长因子受体结合,C型凝集素受体信号传导,白介素1介导的信号传导,通过MHC I类的TAP依赖性抗原处理以及肽抗原的呈现,刺激T细胞受体信号传导,刺激性T细胞受体信号传导以及先天的响应信号通路,信号传输和细胞座的免疫通道富集。高度整合的De-cov-HG被选择构建蛋白质蛋白质相互作用(PPI)网络。结果:使用WalkTrap算法高度互连模块包括模块1(202个节点);从PPI网络检索模块2(126个节点)和模块3(121个节点)模块。MYC,HDAC9,NCOA3,CEBPB,VEGFA,BCL3,SMAD3,SMAD3,SMURF1,KLHL12,CBL,ERBB4和CRKL被鉴定为潜在的药物靶标(PDT),这些药物靶标(PDT)在COV感染后在人类呼吸系统中高度表达。功能项生长因子受体结合,C型凝集素受体信号传导,白介素1介导的信号传导,通过MHC I类的TAP依赖性抗原处理以及肽抗原的呈现,刺激T细胞受体信号传导,刺激性T细胞受体信号传导以及先天的响应信号通路,信号传输和细胞座的免疫通道富集。

抗核纳病毒植物次生代谢产物的潜力...
尽管抗病毒药物开发已经增长,并且疫苗已经可以访问,但仍需要具有成本效益且易于适用的治疗方法来打击Covid-19 [13]。可以口服或通过吸入来施用广谱冠状病毒抑制剂,可能在处理新兴的SARS-COV-2变体方面起着至关重要的作用[13]。这种疗法将对未来的致病性冠状病毒的爆发的准备将是极大的[13]。响应于199的大流行,已经对SARS – COV-2蛋白质和病毒细胞蛋白复合物的结构特性进行了许多研究,以找到治疗性干预措施的潜在靶标[14]。尖峰蛋白,主蛋白酶(MPRO),木瓜样蛋白酶(PLPRO)和RNA-脱纤维RNA聚合酶(RDRP)是最深入研究的药理靶标[14]。通常,针对

人工智能技术的实践及其在对抗中的价值
摘要 当今医学界正处于人工智能时代,人工智能的开发和普及得益于 2019 年冠状病毒病 (COVID-19) 大流行,这为人工智能在分析医疗数据和提供非常准确的结果方面发挥更大影响力提供了空间。这门科学当之无愧地在医护人员中占据了极好和重要的地位,由于其在实际决策中具有巨大的潜力,它已成为医护人员工作中必不可少的要素。由于智能系统能够分析大数据并给出准确的结果,旨在改善公民的健康状况并挽救他们的生命,因此在医疗领域使用智能系统的前景在卫生部门中被视为至关重要。在本文中,重点介绍了一组有关人工智能在医学领域的重要作用的重要信息。此外,该科学如何通过强调一系列调查和分析来应对 SARS‐CoV-2,这些调查和分析在预测病毒传播、追踪感染和通过 COVID-19 患者的胸部 X 光图像诊断病例方面发挥了作用。本文的数据库涵盖了 2020 年至 2021 年之间的 40 多项研究,并研究了利用人工智能技术分析 SARS‐CoV-2 数据的效果。这些研究来自 PubMed、NCBI、谷歌学术、Medrxiv 和其他网站。本文包含大量有关人工智能和 SARS‐CoV-2 的信息。研究结果证实,人工智能在医疗保健领域发挥着重要作用,建议在决策方法中使用其应用。

在计算机分析中,用于重新利用SARS-COV-2的多个靶标的广谱抗病毒药物:一种分子对接和ADMET方法
背景:在严重的急性呼吸综合症冠状病毒-2(SARS-COV-2)引起的第二波COVID-19引起的大流行中,世界遭受了破坏,并导致数百万具有致命毒力潜力的人死亡。与类似的病毒暴发相比,例如严重的急性呼吸综合征冠状病毒(SARS COV)和中东呼吸综合征冠状病毒(MERS COV),Covid-19导致严重的发病率和死亡率。对抗SARS-COV-2感染的各种治疗性干预措施已积极研究,但尚无针对SARS-COV-2病毒具有高抗病毒疗效的特定药物。目前的工作是代表52种广谱抗病毒药物作为抑制SARS-COV-2感染的潜在铅分子的有希望的治疗功效。这些药物已经对几种病毒感染显示了潜在的功效。本文通过使用分子对接方法评估受体 - 配体相互作用来讨论可用广谱抗病毒药物的治疗功效的比较分析。

针对SAR的药物开发的最新进展...
2019年小说冠状病毒(2019-NCOV,后来被称为SARS-COV-2)的突然爆发迅速变成了冠状病毒病的前所未有的2019年冠状病毒大流行(Covid-19)。这次全球医疗保健紧急情况标志着进入新千年后,致命的冠状病毒(COV)进入了人类社会,这使全球医疗体系不堪重负并影响了全球经济。但是,Covid-19的治疗选择仍然非常有限。在病毒生命周期中开发靶向重要蛋白的药物是克服这一困境的一种可行方法。主要蛋白酶(M Pro)在处理COV编码的多蛋白中起主要作用,该多蛋白介导复制转录机制的组装,因此被公认为是理想的抗病毒靶标。在这里,我们总结了针对M Pro的反SARS-COV-2代理发现的最新进展。结合了结构研究,虚拟筛选和实验筛选,已经提出了许多治疗性候选物,包括重新利用的药物和AB始于设计的化合物。科学界的这样的合作努力将加速为Covid-19制定有效治疗的节奏。


2021 年毒药和治疗用品修正案(COVID-19 疫苗)条例
解释性说明 本法规的目的在于: (a) 规定卫生部部长批准可由药剂师供应和施用的 SARS–CoV–2(COVID-19) 疫苗,以及 (b) 授权药剂师以批发方式向授权护士和助产士供应经批准的毒药和限用物质,用于药房疫苗接种。 本法规根据 1966 年《毒药和治疗用品法》制定,包括第 17(1)(a) 和 (c) 条以及第 45C 条(一般法规制定权)。
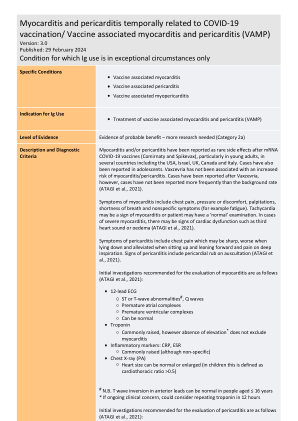
疫苗相关性心肌炎和心包炎 (VAMP
易感人群发生心肌炎的机制尚不清楚,目前已描述了多种潜在机制。免疫系统可能会将疫苗中的 mRNA 检测为抗原,从而激活促炎级联和免疫途径,这些途径可能作为全身反应的一部分在心肌炎的发展中发挥作用。SARS-CoV-2 的刺突蛋白和自身抗原(包括肌球蛋白)之间可能存在分子模拟。心脏反应性自身抗体的产生可能对心脏单核细胞产生功能影响(Bozkurt B et al.,2021)。

冠状病毒 COVID-19:全球各公司、期刊和组织提供的免费文献
心脏 — 心脏病专家和冠状病毒:照顾好自己和彼此,这样我们才能继续照顾患者 BMJ 模拟与技术增强学习 — 通过模拟和技术增强学习,为医疗专业人员准备和应对 2019 年新型冠状病毒 BMJ Open 呼吸研究 — 婴儿急性呼吸道感染对 3 岁儿童肺功能测试的影响 普通精神病学 — 对 COVID-19 疫情中中国人心理困扰的全国性调查 BMJ Open — 韩国中东呼吸综合征冠状病毒 (MERS-CoV) 爆发期间信任对两种不同风险感知作为情感和认知维度的影响 英国眼科学杂志 — 识别可能的早期眼部表现和使用防护眼镜的重要性 欧洲医院药学杂志 — 休斯顿我们有一个问题:冠状病毒 肠道 — SARS-CoV-2 引起的腹泻是 COVID-19 患者的发病症状 肠道 — 腹泻可能被低估:一个缺失的环节2019年新型冠状病毒 BMJ Global Health — MERS-CoV 研究和产品开发的最新路线图 家庭医学与社区健康 — 新型冠状病毒全球爆发中家庭医生面临的挑战和责任 家庭医学与社区健康 — 社区组织在生物危害方面的准备

心肌炎和基于DNA的抗SARS-COV-2疫苗-IJRPR 使用太阳能-IJRPR 动态无线充电站 使用机器学习的疾病预测-IJRPR 自动除颤器对心脏患者的远程监控-IJRPR AI驱动的车辆电池故障检测,监视和预测 开发自动电动汽车-IJRPR 电动两轮车的设计和模拟。 -ijrpr 印度采用电动汽车:评估当前状态和客户看法 来自Makurdi Metropolis选定市场出售的未经糊化牛奶的细菌分离株的细菌学评估。 遗传疾病治疗基因治疗的进步
一名没有心血管危险因素或特定病史的42岁患者,也没有传染病的史。该患者被送入心脏病学部门,用于治疗与接受抗SARS COV 2 DNA疫苗的第一次剂量后四天发生的四天相关的腹部疼痛。患者报告了持续的胸骨后胸痛,在静止和劳累期间发生,躺在左侧而没有任何特定的辐射,从而恶化。这与增加心跳的感觉有关,没有晕厥史或前同步史。