XiaoMi-AI文件搜索系统
World File Search SystemMyc

弥漫性大B细胞淋巴瘤的前线治疗
弥漫性大B细胞淋巴瘤(DLBCL)是非霍奇金淋巴瘤最常见的亚型。尽管这是一种侵略性疾病,但大多数患者对初始治疗做出了很好的反应。标准前线治疗是六个周期的R-Chop化学免疫性疗法,但是阶段和预后特征或危险因素等因素可能会确定缩写是否合适。放射疗法可能是计划治疗过程的一部分,通常是在化学免疫疗法后作为合并。可能需要根据高风险的分子特征(MYC,BCL2和/或BCL6重排),中枢神经系统(CNS)参与诊断或患者在高危次级CNS复发中,可能需要考虑其他化学疗法。在加拿大,没有统一的国家准则来治疗DLBCL,并且存在的省级准则各不相同。有必要由加拿大血液学家支持的基于证据的国家治疗指南,以确保对DLBCL患者的一致治疗采取一致,最佳的方法。来自加拿大各地的一群专家制定了一项国家循证治疗指南,为医疗保健专业人员提供了明确的指南和前线DLBCL管理的最佳实践。根据现有的证据提出了共识建议,提出了当前省级准则的结果。

CLK 激酶抑制剂 1C8 和 GPS167 对上皮-间质转化和抗病毒免疫反应的影响揭示了它们的抗癌潜力
二杂芳酰胺基化合物 1C8 和氨基噻唑酰胺相关化合物 GPS167 可抑制 CLK 激酶,并影响多种癌细胞系的增殖。之前使用 GPS167 进行的化学基因组学筛选表明,与有丝分裂纺锤体组装相关的成分的消耗会改变对 GPS167 的敏感性。在这里,使用 1C8 进行的类似筛选也确定了参与有丝分裂纺锤体组装的成分的影响。因此,用 1C8 和 GPS167 处理的细胞的转录组分析表明,编码有丝分裂纺锤体组装成分的转录物的表达和 RNA 剪接受到影响。通过显示影响有丝分裂纺锤体组装的药物的亚毒性浓度会增加对 GPS167 的敏感性,证实了微管连接的功能相关性。1C8 和 GPS167 影响与肿瘤进展相关的通路中转录本的表达和剪接,包括 MYC 靶标和上皮间质转化 (EMT)。最后,1C8 和 GPS167 改变了参与抗病毒免疫反应的转录本的表达和可变剪接。与此观察结果一致,消耗双链 RNA 传感器 DHX33 可抑制 GPS167 介导的 HCT116 细胞细胞毒性。我们的研究揭示了 1C8 和 GPS167 影响癌细胞增殖以及转移关键过程的分子机制。

对 CDK12 抑制的反应和抵抗......
尽管在弥漫性大 B 细胞淋巴瘤 (DLBCL) 和套细胞淋巴瘤 (MCL) 患者的治疗方面取得了重大进展,但由于耐药性的出现和随后的疾病进展,复发患者的预后仍然很差。迫切需要寻找这些疾病的新靶点和治疗策略。在这里,我们报告 MCL 和 DLBCL 都对转录靶向药物极其敏感,特别是 THZ531,一种细胞周期蛋白依赖性激酶 12 (CDK12) 的共价抑制剂。通过实施药物基因组学和基于细胞的药物筛选,我们发现 THZ531 可抑制致癌转录程序,尤其是 DNA 损伤反应通路、MYC 靶基因和 mTOR-4EBP1-MCL-1 轴,从而有助于体外显著抑制淋巴瘤。我们还从头鉴定和建立了获得性 THZ531 耐药淋巴瘤细胞,这些细胞是由 MEK-ERK 和 PI3K-AKT-mTOR 通路过度激活以及多药耐药性-1 (MDR1) 蛋白上调所致。值得注意的是,EZH2 抑制剂通过竞争性抑制 MDR1 逆转了对 THZ531 的耐药性,并与 THZ531 联合使用,在体外协同抑制了 MCL 和 DLBCL 的生长。我们的研究表明,CDK12 抑制剂单独使用或与 EZH2 抑制剂联合使用,有望成为难治性 DLBCL 和 MCL 的新型有效治疗方法。

最初发表于:Saikumar Lakshmi, Priya;Oduor, Cliff I;Forconi, Catherine S;M'Bana, Viriato;Bly, Courtney;Gerstein, Rachel M;Otieno, Ju
地方性伯基特淋巴瘤 (BL) 是撒哈拉以南非洲的一种儿童癌症,其特征是爱泼斯坦-巴尔病毒和疟疾相关的异常 B 细胞活化和 MYC 染色体易位。常规化疗后的存活率徘徊在 50% 左右;因此,需要临床相关模型来测试其他疗法。因此,我们建立了五种患者来源的 BL 肿瘤细胞系和相应的 NSG-BL 化身小鼠模型。转录组学证实,我们的 BL 系从患者肿瘤到 NSG-BL 肿瘤都保持了保真度。然而,我们发现 NSG-BL 化身之间的肿瘤生长和存活率以及爱泼斯坦-巴尔病毒蛋白表达模式存在显著差异。我们测试了利妥昔单抗的反应性,发现一种 NSG-BL 模型表现出直接敏感性,其特点是凋亡基因表达与未折叠蛋白反应和 mTOR 促生存途径相平衡。在利妥昔单抗无反应的肿瘤中,我们观察到 IFN-α 特征,这由 IRF7 和 ISG15 的表达证实。我们的结果表明患者间肿瘤存在显著的差异和异质性,并且当代患者衍生的 BL 细胞系和 NSG-BL 化身是指导新治疗策略和改善这些儿童预后的可行工具。

对人类原代自然杀伤细胞的基因操作......
淋巴结外自然杀伤 (NK)/T 细胞淋巴瘤,鼻型 (ENKTCL) 是一种高度侵袭性的淋巴瘤,其中肿瘤抑制基因 PRDM1 经常丢失或失活。我们采用了两种不同的 CRISPR/Cas9 方法来生成 PRDM1 -/- 原代 NK 细胞,以研究该基因在 NK 细胞稳态中的作用。与野生型相比,PRDM1 -/- NK 细胞的克隆效率显著提高、增殖率更高、凋亡更少。基因表达谱显示,在 PRDM1 -/- NK 细胞中,与增殖、细胞周期、MYC、MYB 和 TCR/NK 信号相关的通路显著富集,但与正常细胞功能(包括细胞毒功能)相关的通路被下调,这表明 PRDM1 的缺失使 NK 细胞转向增殖和存活,而不是发挥其正常功能。我们还能够进一步修改 PRDM1 缺失的克隆,以引入 ENKTCL 中常见的肿瘤抑制基因(如 TP53、DDX3X 和 PTPN6)的杂合缺失。我们建立了体外模型来阐明 PRDM1 介导其对 NK 细胞的稳态控制的主要途径。这种方法可以应用于研究淋巴瘤发病机制中的其他相关遗传病变和致癌协同作用。

抗有丝分裂药物 PTC-028 和 PTC596 在多发性骨髓瘤临床前模型中表现出强效活性,但对 BMI-1 作为必需肿瘤基因的作用提出了质疑
多发性骨髓瘤 (MM) 治疗的未来进展需要表征该疾病的主要驱动因素,并采用新颖的创新方法来解决这些弱点。本研究重点关注一种新型药物 BMI-1 调节剂在 MM 中的临床前评估。我们在一系列体外和体内模型(包括耐药性和基质支持模型)中证明了 PTC-028 和 PTC596 的强效活性。用 PTC-028 和 PTC596 治疗 MM 细胞会下调 BMI-1 蛋白水平,发现这与药物活性相关。令人惊讶的是,BMI-1 对 BMI-1 调节剂的活性和 MM 细胞生长是可有可无的。我们的数据表明,有丝分裂停滞伴有髓细胞白血病-1 (MCL-1) 丢失是关键的抗 MM 机制,并揭示了由于 BMI-1 调节剂治疗导致 MYC 和 AKT 信号传导活性受损。此外,我们在 5TGM.1 体内模型中观察到 PTC596 治疗后 MM 完全消除,并将表观遗传化合物和 B 细胞白血病/淋巴瘤 2 同源域 3 (BH3) 模拟物定义为有希望的组合伙伴。这些结果对 BMI-1 作为必需 MM 基因的假定作用提出了质疑,并证实 BMI-1 调节剂是有效的抗有丝分裂剂,具有令人鼓舞的临床前活性,支持其快速转化为临床试验。
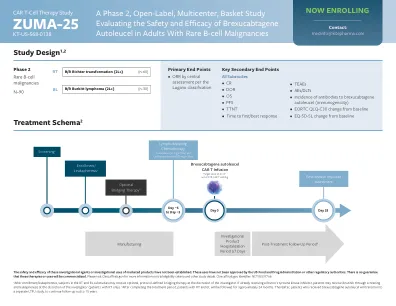
Zuma -25 -Gilead and Kite肿瘤学|医疗事务
aes,不利事件; ANC,绝对中性粒细胞计数; BCL,B细胞淋巴瘤; BL,伯基特淋巴瘤;汽车T,嵌合抗原受体T细胞疗法; CD4/19,分化簇4/19; CLL,慢性淋巴细胞性白血病;中枢神经系统,中枢神经系统; CR,完全响应; CSF,脑脊液; DLBCL,弥漫性大B细胞淋巴瘤; DL,十分位; DLT,剂量限制毒性; DOR,响应持续时间; ECOG PS,东方合作肿瘤学组绩效状况; EORTC QLQ-C30,欧洲研究和治疗癌症质量的生活问卷的组织; 30; EQ-5D-5L,欧洲生活质量五个维度五个级别的规模; GVHD,移植-VS-Host-疾病; HBV,乙型肝炎病毒; HCV,丙型肝炎病毒; HGBCL,高级B细胞淋巴瘤;超瓦德,超分级环磷酰胺,长春新碱,阿霉素和地塞米松; LTFU,长期随访; MYC,细胞周期进入和增生代谢的主要调节剂; NHL,非霍奇金淋巴瘤; HIV,人类免疫缺陷病毒; ORR,客观响应率; OS,整体生存; PFS,无进展的生存; r/r,复发或难治; RT,Richter转型; SCT,干细胞移植; TEAE,治疗急性不良事件; TTNT,下一次治疗的时间。

基于血液的神经内分泌肿瘤的下一代测序分析
结果:320 例 NEN 患者中,182 例(57%)为男性,中位年龄为 63 岁(范围:8-93 岁)。肿瘤类型包括胰腺 NET(N = 165,52%)、胃肠道 NEC(N = 52,16%)、大细胞肺 NEC(N = 21,7%)、鼻咽 NEC(N = 16,5%)和未另作规定的 NEC/NET(N = 64,20%)。对 338 份血浆样本进行了 ctDNA NGS 检测;14 例患者进行了两次检测,2 例患者进行了三次检测。在 280 例(87.5%)样本中发现了基因组变异,在排除意义不明确的变异 (VUS) 和同义突变后,共识别出 1,012 处变异。在 280 个发生改变的样本中,TP53 相关基因发生改变最多(N = 145, 52%),其次是 KRAS(N = 61, 22%)、EGFR(N = 33, 12%)、PIK3CA(N = 30, 11%)、BRAF(N = 28, 10%)、MYC(N = 28, 10%)、CCNE1(N = 28, 10%)、CDK6(N = 22, 8%)、RB1(N = 19, 7%)、NF1(N = 19, 7%)、MET(N = 19, 7%)、FGFR1(N = 19, 7%)、APC(N = 19, 7%)、ERBB2(N = 16, 6%)和 PTEN(N = 14, 5%)。

多发性骨髓瘤:BET蛋白水解靶向嵌合分子与CDK9抑制剂联合治疗
细胞周期蛋白依赖性激酶 9 (CDK9) 与溴结构域和末端外结构域 (BET) 蛋白结合,通过磷酸化 RNAP II C 末端结构域的丝氨酸 2 来促进转录延长。我们在体外研究了选择性 CDK9 抑制剂 (AZD 4573 和 MC180295) 对人类多发性骨髓瘤细胞的治疗潜力。与对照细胞相比,多发性骨髓瘤 (MM) 细胞系中 CDK9 的短发夹 RNA 沉默降低了细胞活力,表明 MM 细胞对 CDK9 的依赖性。为了探索与 CDK9 抑制剂的协同作用,添加了蛋白水解靶向嵌合分子 (PROTAC) ARV 825。后一种药物导致 BET 蛋白泛素化,从而导致其快速高效降解。与单独使用任何一种药物相比,ARV 825 和 AZD 4573 联合治疗 MM 细胞可显著降低其 BRD 2、BRD 4、MYC 和磷酸化 RNA pol II 的蛋白质表达。联合治疗在体外和体内协同抑制多发性骨髓瘤细胞,且体重减轻不明显。与单独使用一种药物相比,该组合还导致低剂量的凋亡细胞显著增加。总之,我们的研究首次表明 BET PROTAC(ARV 825)和 AZD 4573(CDK9 抑制剂)的组合对 MM 细胞有效。
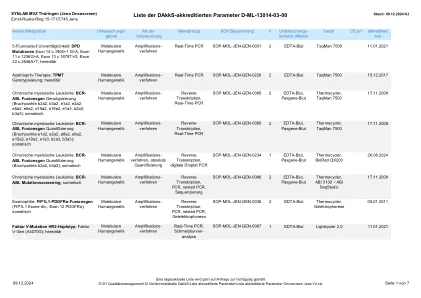
认可参数列表 Oncoscreen Jena v5.xlsx
Oncomine Comprehensive Assay v3 DNA 组:AKT1、AKT2、AKT3、ALK、AR、ARAF、ARID1A、ATM、ATR、ATRX、AXL、BAP1、BRAF、BRCA1、BRCA2、BTK、CBL、CCND1、CCND2、CCND3、CCNE1、CDK12、CDK2、CDK4、CDK6、CDKN1B、CDKN2A、CDKN2B、CHEK1、CHEK2、CREBBP、CSF1R、CTNNB1、DDR2、EGFR、ERBB2、ERBB3、ERBB4、ERCC2、ESR1、EZH2、FANCA、FANCD2、FANCI、FBXW7、FGF19、FGF3、FGFR1、FGFR2、FGFR3、FGFR4、FLT3、 FOXL2、GATA2、GNA11、GNAQ、GNAS、H3-3A、HIST1H1E、HNF1A、HRAS、IDH1、IDH2、IGF1R、JAK1、JAK2、JAK3、KDR、KIT、KNSTRN、KRAS、MAGOH、MAP2K1、MAP2K2、MAP2K4、MAPK1、MAX、MDM2、 MDM4、MED12、MET、MLH1、MRE11A、MSH2、MSH6、MTOR、MYC、MYCL、MYCN、MYD88、NBN、NF1、NF2、NFE2L2、NOTCH1、NOTCH2、NOTCH3、NRAS、NTRK1、NTRK2、NTRK3、PALB2、PDGFRA、PDGFRB、PIK3CA、 PIK3CB, PIK3R1、PMS2、POLE、PPARG、PPP2R1A、PTCH1、PTEN、PTPN11、RAC1、RAD50、RAD51、RAD51B、RAD51C、RAD51D、RAF1、RB1、RET、RHEB、RHOA、RICTOR、RNF43、ROS1、SETD2、SF3B1、SLX4、SMAD4、SMARCA4、SMARCB1、SMO、SPOP、SRC、STAT3、STK11、TERT、TOP1、TP53、TSC1、TSC2、U2AF1、XPO1