XiaoMi-AI文件搜索系统
World File Search SystemPTMS

UC Santa Barbara
引言病理tau蛋白在神经退行性疾病中积累并形成了大脑中的聚集体,这些疾病被归类为tauopathies,包括阿尔茨海默氏病(AD),Pick's Disaise,Corticobasal Denneration(CBD)和进行性性超额外核Palsy Palsy(PSP)。aberant tau翻译后修饰(PTMS)在大脑中发现的可以改变tau蛋白功能,促进tau聚集并触发细胞中的毒性(1,2)。在人脑中积累的病理修饰的TAU的程度与疾病中痴呆症的严重程度相关(3,4)。tau主要在健康神经元的轴突中进行定位,而在AD大脑中的突触中发现了大量的病理学tau(5-7)。合成功能障碍是在神经变性之前的tauopathy小鼠模型中最早的病理生理变化之一,并且与认知障碍的开始相吻合(8-10)。长期增强(LTP)是海马突触时的重要可塑性机制,它是新记忆形成的基础,而LTP的抑制作用被TAU抑制,而Tau的突变引起家族性额额额痴呆(FTD)(FTD)(FTD)(8、10、11)(8、10、11),以及几种伴随的patagencied hypersocied hypercried the tau tau tau(9) Tau(12,13)和Tau低聚物(14,15)。
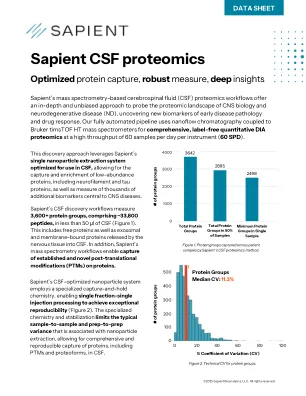
CSF蛋白质组学服务|数据表 - 智商生物
在来自ND和匹配的对照组患者的CSF的病例对照研究中,发现ND患者的CSF在CSF中显着增加了神经元损伤的关键生物标志物,包括神经丝链轻链蛋白和TAU蛋白(图6A)。重要的是,在ND患者的CSF中发现了数千种其他蛋白质(包括蛋白质成型和PTM)的其他蛋白质差异存在,揭示了潜在的新ND生物标志物或药物靶标(图6B)。

蛋白质生产的无细胞未来
尽管自发现遗传密码(9)以来,无细胞的系统就已经存在,但其产量低,无法正确实施PTM,并且缺乏可伸缩性限制了其主要用于研究实验室的使用。通过克服这些广泛采用的这些关键障碍,像爱丽丝这样的多功能细胞系统开始改变蛋白质生产中的当前范式。实现可伸缩性是朝着这一目标迈出的重要一步。快速合成能力,以及它们大规模表达广泛功能蛋白的能力,无细胞的系统设置为重新定义生物制造。

表征与液相色谱质谱法(LC-MS)的肽相关病毒(AAV)衣壳蛋白的基于液相色谱(LC-MS)的肽映射和翻译后修饰分析(PTMS)
肽映射样品制备:AAV8参考材料在2x10 13 Vg/ml的浓度下包含20μl的总体积。这导致消化的估计总蛋白浓度为0.12μg/μL,总蛋白质为2.4μg。将AAV样品在6 m尿素中变性,在80℃以1 mm DTT变性30分钟,然后用15 mm iodoacetamide烷基化在黑暗中的室温下在室温下30分钟。将还原和烷基化的样品冷却至室温,并用3次同等体积的缓冲液(50 mM Tris-HCl和1 mm CaCl 2 [pH 7.5])稀释,将尿素浓度降低至<2M。然后将样品降低到<2M。然后用0.4 µ µGGGRYPSIN或CHYMOTRYPESIN或CHYMOTRYPSIN或CHYMOTRYPRYRYPERSIN或CHYMOTRYPRYRYPRYRYPRYRYPRYRYSIL逐夜消化。通过将甲酸添加到最终浓度的10%中终止消化,并将样品直接注入LCMS-9050进行分析。

T细胞功能核心的代谢和表观遗传学
t细胞子群根据其功能和周围的微环境适应并重新加油其新陈代谢。幼稚的T细胞依赖于以养分需求低的方式为特征的线粒体代谢途径,而效应T细胞诱导动力学途径更快,以产生增殖和细胞因子产生所需的生物量和能量。最近发现的概念是代谢的改变也会影响T细胞的表观遗传学。在这篇综述中,我们讨论了T细胞代谢和表观遗传变化之间的联系,例如组蛋白后翻译后修饰(PTMS)和DNA甲基化,以及代谢酶和分子的“超代谢”作用。这些发现集体指出了一组新的潜在治疗靶标,用于治疗T细胞依赖性自身免疫性疾病和癌症。

预测基于 CRISPR-Cas9 的表观基因组编辑的效果
摘要表观遗传调控协调哺乳动物转录,但它们之间的功能联系仍然难以捉摸。为了解决这个问题,我们使用来自 13 种 ENCODE 细胞类型的表观基因组和转录组数据来训练机器学习模型,以预测组蛋白翻译后修饰 (PTM) 的基因表达,对于大多数细胞类型,实现了 ∼0.70 −0.79 的转录组范围相关性。我们的模型重现了组蛋白 PTM 和表达模式之间的已知关联,包括预测转录起始位点 (TSS) 附近的组蛋白亚基 H3 赖氨酸残基 27 (H3K27ac) 的乙酰化会显著提高表达水平。为了通过实验验证这一预测,并研究 H3K27ac 的天然沉积与人工沉积对表达的影响,我们将合成的 dCas9-p300 组蛋白乙酰转移酶系统应用于 HEK293T 细胞系中的 8 个基因和 K562 细胞系中的 5 个基因。此外,为了便于建立模型,我们执行 MNase-seq 来绘制 HEK293T 中全基因组核小体占有水平。我们观察到,我们的模型在准确排序基因对 dCas9-p300 系统的相对倍数变化方面表现良好;然而,与根据其天然表观遗传特征预测跨细胞类型的表达相比,它们对单个基因内倍数变化进行排序的能力明显减弱。我们的研究结果强调,我们需要更全面的基因组规模表观基因组编辑数据集,更好地理解表观基因组编辑工具所做的实际修改,以及改进因果模型,以便更好地从内源性细胞测量转移到扰动实验。这些改进将共同促进理解和可预测地控制动态人类表观基因组的能力,以及对人类健康的影响。

定性/定量蛋白质组学相互作用分析...
蛋白质组学是所有蛋白质的蛋白质,以及它们的细胞,组织,体液(例如血清/血浆或脑脊液)或各个基因组在给定时间点表达的整个生物体中的转化后修饰(PTM)。这可以扩展为包括对定义的刺激或疾病状态的蛋白质(和PTM)水平变化的深入定量分析。这些目标通常是通过样品分离技术(例如色谱方法和超高分辨率质谱法)(Nanolc-MS/ MS)的组合来实现的。蛋白质组学现在是一种标准的分析,非常有力的研究技术,可以在RWTH AACHEN的生物医学领域(生物化学,细胞生物学,分子医学,系统生物学等)中用于基本上进行的所有研究。)。

免疫疗法和癌症:多摩学观点
免疫疗法(IT)代表了癌症治疗方面的显着成就[1]。肿瘤免疫疗法通过重新启动肿瘤免疫周期并恢复人体的天然抗肿瘤免疫反应来起作用[2]。目前,至少有四种主要的免疫疗法策略,其中包括免疫检查点抑制剂(ICIS),例如程序性细胞死亡蛋白1(PD-1)和细胞毒性T淋巴细胞抗原4(CTLA-4),嵌合抗原受体T-Cell受体T-Cell Therof actapy,Tumory pacocine and Tumory pacocines,thmory和Peripications and Peripatications和Peripaticationcation。尽管这些疗法已广泛成功,但增强了临床肿瘤结局[2],但并非所有患者都从中受益[1]。因此,对于从免疫疗法中获得最多的筛查至关重要[2]。肿瘤异质性可能是由于遗传,表观遗传和转录修饰等多种因素而产生较低治疗疗效的原因。蛋白质表达变化;以及代谢谱的变化[3]。最近,人们非常关注翻译后修饰(PTMS),这些变化是对单个氨基酸的小变化,例如糖基化,乙酰化,乙酰化,磷酸化,棕榈酰化和泛素化或泛素化或去泛素化。已经发现这些PTM具有改变蛋白质与其他分子的功能,形状,平衡和相互作用的能力。此外,最近的研究表明,PD-1和程序性细胞死亡配体1(PD-L1)的表达水平可以受到表观遗传,转录和转录后系统的调节,从而影响肿瘤免疫[4,5]。在这种情况下,多词的方法结合了基因组学,转录组学,蛋白质组学,代谢组学,放射组学和免疫学,有助于揭示肿瘤中存在的各种层次,并探索蛋白质内的双重性,并探索蛋白质丰富的蛋白质,代表性地表现出跨性别的细胞表达,摩尔纳的形式和基因型的摩擦性,基因构图,基因构想,基因构图,基因范围,基因范围,莫尔纳(MRNA)的概述,莫尔纳(MRNA)的概述,莫尔娜癌症以及肿瘤 - 免疫间隔机制,鉴定出新的潜在生物标志物和免疫疗法靶标,并促进与免疫疗法相关的独特分子特征的鉴定

胰腺癌中的 SUMO 通路
迫切需要为胰腺导管腺癌 (PDAC) 患者开发新的治疗策略。然而,尽管在 PDAC 的组织病理学和分子亚型方面做出了各种努力,但尚未建立新的靶向或特异性疗法。具有泛素样蛋白的翻译后修饰 (PTM),包括小泛素样修饰物 (SUMO),介导许多有助于癌细胞适应性和存活的过程。SUMO 化对转录控制、DNA 修复途径、有丝分裂进展和致癌信号传导的贡献已被描述。本文我们回顾了 PDAC 中 SUMO 通路的功能,特别关注其与以高 MYC 活性为特征的侵袭性疾病亚型的联系,并讨论了正在开发的用于精准 PDAC 疗法的 SUMO 化抑制剂。
