XiaoMi-AI文件搜索系统
World File Search SystemRAT

消费绵羊和牛奶对大鼠脑脂肪酸和磷脂成分的影响
在两个28天的喂养实验中研究了羊奶和牛奶对大鼠脑脂质组成的影响。使用乙醇 - 己烷提取大鼠脑的总脂质,并使用带有火焰电离检测的气相色谱法(GC-FID)和磷酸磷酸化的脂肪酸和磷脂含量(磷脂)和磷-31核磁共振(31 P NMR)。此外,使用衰减的总反射率傅立叶变换红外和傅立叶变换拉曼规格Troscopicy分析了冷冻干燥的合并样品,并使用多变量方法进行了分析。与研究1中的羊奶处理的组相比,在牛奶组中发现了C18:2更高的C18:2含量。在研究第二,与对照低Ca/p组相比,绵羊牛奶处理的组中的C16:0含量显着(P <0.05)。在光谱分析中没有观察到显着(p> 0.05)。可以得出结论,喂给大鼠28天的绵羊和牛奶对脑脂肪组的影响很低。

在坐骨神经损伤大鼠模型中,尿苷的再生作用中表观遗传机制的介导
在横切损伤中,外周神经的退化变化发生在损伤的两侧,从而观察到部分或完全的感觉/运动损失(8,22,43,47)。在周围神经损伤后,近端段发生退化性变化,远端段发生沃勒(Wallerian)变性(30)。病理生理的变化,例如凋亡,氧化应激,炎症,细胞外基质的破坏以及其他几个事件可能会使周围神经损伤(PNI)的损害程度恶化(29,48,49,49,52);但是,这些复杂的过程在每个阶段都可以破坏以防止伤害后再生。尽管已经开发了针对这些过程的几种手术和医学方法,但可以保证PNI的功能恢复的治疗方法尚未发现(8,30,43,47)。

小鼠和大鼠排卵前卵泡中 LH 受体与 NPR2 鸟苷酸环化酶之间的信号传导
• LH 诱导的 NPR2 去磷酸化可能不是由 PPP 家族磷酸酶活性的变化介导的。 • GSK3A/B 是 NPR2 调节位点的候选激酶。 • LH/PKA 信号传导使 GSK3A/B 上的抑制位点磷酸化。 • GSK3 的抑制剂会导致 NPR2 去磷酸化和 NEBD。 未来方向 • 使用 GSK3A(全局);GSK3B(颗粒特异性)敲除小鼠来测试 GSK3 是否是维持 NPR2 磷酸化所必需的。 • 使用 GSK3A-S21A/S21A;GSK3B-S9A/S9A 12 突变小鼠来测试 GSK3A/B 磷酸化是否是 LH 诱导的 NPR2 去磷酸化和减数分裂恢复所必需的。

糖尿病ZDSD大鼠模型中肠道微生物群的纵向表征和寡聚果糖的治疗潜力
摘要:2型糖尿病(T2D)的复杂发展为研究动物模型中疾病的进展和治疗带来了挑战。新开发的糖尿病大鼠模型,Zucker糖尿病Sprague Dawley(ZDSD)大鼠,与人类T2D的进展紧密相似。在这里,我们检查了雄性ZDSD大鼠T2D和肠道菌群中相关的变化的进展,并测试该模型是否可用于检查潜在疗法的效率,例如益生元,特定寡寡素化的,靶向了gut microbobiota。体重,肥胖,喂养/空腹血糖和胰岛素。葡萄糖和胰岛素耐受性测试,并使用16S rRNA基因测序在8、16和24周龄进行短链脂肪酸和微生物群分析时收集的粪便。在24周结束时,一半的大鼠补充了10%的寡果糖,并重复测试。我们观察到通过恶化的胰岛素和葡萄糖耐受性,从健康/非糖尿病患者到糖尿病前期和公开糖尿病态的过渡,进食/禁食葡萄糖的显着增加,然后显着减少循环胰岛素。与健康和糖尿病前期相比,在公开糖尿病状态下,乙酸和丙酸酯水平显着增加。微生物群分析表明,与糖尿病前和糖尿病态相比,健康型和β多样性的变化以及健康属的变化以及特定细菌属的变化发生了变化。寡聚果糖治疗改善了葡萄糖耐受性,并在晚期糖尿病期间改变了ZDSD大鼠的盲肠菌群。这些发现强调了ZDSD大鼠作为T2D模型的转化潜力,并突出了可能影响疾病发展或作为T2D的生物标志物的潜在肠道细菌。此外,寡果糖处理能够中度改善葡萄糖稳态。

一个极化编码的光子到旋转界面
摘要。心力衰竭和骨骼肌弱是糖基因论11型的主要临床特征,这是由酸A-葡萄糖苷酶缺乏引起的溶酶体储存障碍。在我们的研究中,我们已经在大鼠心脏灌注灌注系统中调查了酸A-葡萄糖苷酶是否可以从血管系统中吸收到心脏病中。将大鼠心脏用含有含磷酸盐的甘露糖含有甘露糖的含酸A-葡萄糖苷酶灌注,从Bovine睾丸纯化时,获得了3至4倍的酶活性。灌注含有含有甘露糖的6-磷酸盐识别标记物的人胎盘酸A-葡萄糖酶没有这种作用。通过免疫印迹证明了牛睾丸酸A-葡萄糖苷酶在心脏组织中的存在。免疫细胞化学为摄取心肌细胞溶酶体的外源性酶提供了证据。讨论了这些发现与I1型糖原病中酶治疗的相关性。(Pe-Diatr Res 28:344-347,1990)

自闭症特异性母体自身抗体暴露的大鼠模型中行为、大脑结构和神经代谢发生改变
母体免疫失调是自闭症谱系障碍 (ASD) 的产前风险因素。重要的是,炎症和代谢压力之间存在临床相关联系,可导致异常的细胞因子信号传导和自身免疫。在这项研究中,我们研究了母体自身抗体 (aAbs) 破坏代谢信号传导并诱导暴露后代大脑神经解剖学变化的可能性。为此,我们根据母体自身抗体相关 ASD (MAR-ASD) 的临床现象开发了大鼠母体 aAb 暴露模型。在确认大鼠母体产生 aAb 并将抗原特异性免疫球蛋白 G (IgG) 转移到后代后,我们纵向评估了后代行为和大脑结构。当允许 MAR-ASD 大鼠后代与新伴侣自由互动时,幼崽超声波发声减少,社交游戏行为明显减少。此外,在另一组动物中,在出生后第 30 天 (PND30) 和 PND70 天进行的纵向体内结构磁共振成像 (sMRI) 显示,总体和局部脑容量存在性别差异。在 MAR-ASD 后代中,不同区域的治疗特定影响似乎集中在中脑和小脑结构上。同时,收集体内 1 H 磁共振波谱 (1 H-MRS) 数据以检查内侧前额叶皮质中的大脑代谢物水平。结果表明,与对照动物相比,MAR-ASD 后代的胆碱化合物和谷胱甘肽水平降低,同时牛磺酸水平升高。总体而言,我们发现暴露于 MAR-ASD aAbs 的大鼠表现出行为、大脑结构和神经代谢的改变;让人联想到在临床 ASD 中观察到的发现。

大鼠脑转录组:从婴儿期到衰老和散发性阿尔茨海默病样病理
摘要:有人提出,成人大脑的功能特征(所有这些都是在生命早期形成的)可能会影响大脑对阿尔茨海默病 (AD) 的易感性。我们之前对衰老加速的 OXYS 大鼠(一种散发性 AD 模型)的研究结果支持这一假设。在这里,为了阐明大脑成熟过程中出现的异常的分子遗传性质,我们分析了 OXYS 大鼠和 Wistar(对照)大鼠在大脑成熟的关键时期(P3 和 P10 岁;P:出生后天数)的前额皮质 (PFC) 和海马的转录组(RNA-seq 数据)。我们在两个大脑结构中发现了 1000 多个差异表达基因;功能分析表明神经元接触形成效率降低,这大概主要是由于线粒体功能缺陷所致。接下来,我们比较了从婴儿期到 AD 样病变进展阶段(共五个年龄段)大鼠 PFC 和海马中差异表达的基因。三种基因( Thoc3 、 Exosc8 和 Smpd4 )在整个生命周期中均在 OXYS 大鼠的两个脑区中表现出过度表达。因此,婴儿期 OXYS 大鼠脑中神经网络形成效率的降低可能是导致其出现 AD 样病变的原因。

疫苗介导机制控制大鼠共培养系统中土拉弗朗西斯菌 SCHU S4 的生长
摘要:土拉弗朗西斯菌可引起严重的土拉菌病。本研究的目的是通过研究两种候选疫苗在大鼠和小鼠模型中提供不同程度保护的免疫反应来确定大鼠共培养模型中保护的相关性。免疫反应的特征是使用来自幼稚或活疫苗株 (LVS) 或 ∆ clpB / ∆ wbtC 免疫的 Fischer 344 大鼠的脾细胞作为效应物,以及感染高毒性菌株 SCHU S4 的骨髓来源的巨噬细胞。引发复杂的免疫反应,导致细胞因子分泌、一氧化氮产生和对细胞内细菌生长的有效控制。添加 LVS 免疫脾细胞比 ∆ clpB / ∆ wbtC 脾细胞对细菌生长的控制明显更好。这反映了候选疫苗在大鼠模型中的有效性。与 LVS 免疫大鼠的脾细胞相比,与 ∆ clpB / ∆ wbtC 脾细胞共培养的 IFN-γ、TNF、fractalkine、IL-2 和亚硝酸盐水平较低。发现一氧化氮与保护作用相关,因为一氧化氮水平与保护程度呈负相关,并且抑制一氧化氮产生会完全逆转 SCHU S4 的生长抑制。总体而言,结果表明,与大鼠衍生细胞共培养试验是确定针对 F. tularensis 高毒性菌株保护作用相关因素的合适模型
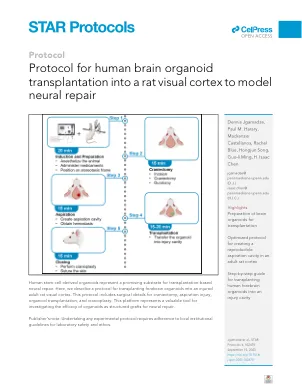
人脑机形的人体器官移植到大鼠视觉皮层中,以建模神经修复
以下方案概述了将人类前脑器官移植到年轻成年男性长埃文斯大鼠的视觉皮层中的特定步骤。使用先前发表的协议生成了本协议中描述的前脑类器官。2,该方法已通过源自诱导的多能干细胞和胚胎干细胞系的器官成功实现。有关此协议尝试使用的单元线的更多详细信息,请参阅我们以前的研究。1,3我们打算主要用于将皮质器官移植到啮齿动物皮质中,但可以适应其他区域身份的器官向皮层移植。这种损伤修复模型提供了增强的转化重要性,从而能够检查活生物体中脑器官的结构和功能属性。此外,它可以作为将神经移植工作扩展到视觉系统以外的大脑区域的基础。

在自闭症特异性母体自身抗体暴露的大鼠模型中改变行为,大脑结构和神经代谢物
母体免疫失调是自闭症谱系障碍(ASD)的产前危险因素。重要的是,炎症和代谢应激之间存在临床相关的联系,这可能导致细胞因子信号传导和自身免疫性异常。在这项研究中,我们检查了孕产妇自身抗体(AABS)破坏代谢信号传导并诱导暴露后代大脑中神经解剖学变化的潜力。为了实现这一目标,我们基于母体自身抗体相关的ASD(MAR-ASD)的临床现象开发了大鼠母体AAB暴露模型。确认大鼠大坝和特异性免疫球蛋白G(IgG)转移到后代后,我们纵向评估了后代行为和大脑结构。mar-asd老鼠后代在允许与新型伴侣自由互动时,表现出幼犬超声发声的减少,并且在社交行为中表现出明显的定义。此外,在产后第30天(PND30)和PND70在单独的动物中进行的纵向体内结构磁共振成像(SMRI)揭示了性别特异性差异。按区域划分的治疗特异性作用似乎在Mar-Asd后代的中脑和小脑结构上汇聚。同时收集了体内1小时磁共振光谱(1 H-MRS)数据,以检查内侧前额叶皮层中的脑代谢物水平。结果表明,与对照动物相比,含胆碱化合物和谷胱甘肽的水平显示出含胆碱化合物和谷胱甘肽的水平降低。总体而言,我们发现暴露于MAR-ASD AAB的大鼠行为,大脑结构和神经代谢物的改变。让人联想到在临床ASD中观察到的发现。