XiaoMi-AI文件搜索系统
World File Search SystemTREM2

DNA 甲基化是 STX6 和其他额颞叶变性遗传风险相关基因位点失调的一个因素
MAPT cg01934064 17 44064242 船体搁板 -0.14 0.024 MAPT cg15323584 17 44022846 5'UTR 搁板 0.11 0.009 MAPT cg17569492 17 44026659 5'UTR 岛 0.09 0.019 MAPT cg12727978 17 44075500 船体露天海域 0.08 0.009 TREM2 cg02828883 6 41131823 TSS1500 露天海域 0.08 0.005 TIA1 cg14434028 2 70452453 船体露天海域 0.08 0.036 TIA1 cg13119546 2 70444039 身体 opensea 0.05 0.041 RUNX2 cg16181497 6 45409732 身体 opensea -0.07 0.042 RUNX2 cg12755953 6 45430813 身体 opensea 0.06 0.039 RUNX2 cg04110902 6 45500999 身体 opensea 0.05 0.038 GRN cg06800040 17 42427647 身体 shelf 0.07 0.022 FTLD1m 按亚型分类:TDP Type A C9orf72 vs CTRL MAPT cg15323584 17 44022846 5'UTR shelf 0.17 0.002 MAPT cg12727978 17 44075500 船体 开海 0.15 0.001 MAPT cg17569492 17 44026659 5'UTR 岛 0.1 0.032 MAPT cg19276540 17 44060353 船体 岛 0.08 0.035 RUNX2 cg12041069 6 45341222 船体 搁板 0.15 0.04 RUNX2 cg17636752 6 45391973 船体 岸 0.09 0.036 RUNX2 cg12755953 6 45430813 船体 开海 0.08 0.026 TIA1 cg14434028 2 70452453 身体 开放海 0.13 0.011 TIA1 cg13119546 2 70444039 身体 开放海 0.06 0.047 TIA1 cg15836561 2 70442511 ExonBnd 开放海 0.06 0.028 TBK1 cg23175599 12 64848891 5'UTR 架 0.1 0.026 TREM2 cg02828883 6 41131823 TSS1500 开放海 0.09 0.017 CCNF cg26647200 16 2482775 身体 架 0.09 0.022 GRN cg06800040 17 42427647 车身搁板 0.08 0.031 GRN cg12837296 17 42426483 5'UTR 开海 0.07 0.033 GRN cg23570245 17 42426011 5'UTR 开海 0.06 0.048 GRN cg08491241 17 42421960 TSS1500 开海 0.06 0.05 SQSTM1 cg05578452 5 179255653 车身开海 0.07 0.005 SQSTM1 cg09046399 5 179264098 3'UTR 开海 0.06 0.025 FTLD1m 亚型:TDP C 型 vs CTRL MAPT cg01934064 17 44064242 船体架 -0.16 0.016 MAPT cg17569492 17 44026659 5'UTR 岛 0.08 0.045 MAPT cg26979107 17 44061355 船体岸 0.06 0.016 MAPT cg22635938 17 44039549 5'UTR 公海 -0.06 0.012 MAPT cg01582587 17 44036817 5'UTR 公海 0.05 0.022 TBK1 cg09999583 12 64878162 船体公海-0.1 0.029 TREM2 cg02828883 6 41131823 TSS1500 公海 0.08 0.009

MACC1调节LGR5以促进结直肠癌的癌症干细胞特性
从HIV-1 + 2,乙型肝炎和丙型肝炎中分离出外周血单核细胞(PBMC)。pBMC,包括人类基因Oct3/4,Sox2,c-Myc和klf4的矢量。HIPSC线BIHI292-A源自单个菌落,并在E8培养基中保持未分化的HIPSC的典型形态(图1 a)。通过PCR确认缺乏仙台病毒载体(Suppl。图1 a)。BIHI292-A HIPSCS ECT3/4,SSEA-4,NANOG和TRA-1 - 60作为未分化HIPSC状态的典型标记,如使用免疫细胞化学所示(图1 b)。进一步的流式细胞仪证实了OCT3/4,SSEA-4,NANOG和TRA-1-60表达在超过96%的BIHI292-A HIPSC中的SSEA-4,Nanog和Tra-1-60表达中的干性标记表达(图。1 c)。g带核分型在GTG上进行(使用GIEMSA的胰蛋白酶G带)进行染色的中期染色体,并揭示了正常的雌性Karyo 46型,XX(图1 D)。 单核苷酸多态性分析表明,与患者的PBMC相比,BIHI292-A HIPSC线没有任何拷贝数变量> 2 Mb> 2 MB> 5 MB(表1)。 短串联重复(STR)分析的结果表明,BIHI292-A细胞系和患者的PBMC的遗传认同是相同的(表1)。 Sanger测序证实了两个Exon 2中的两个Trem2杂合突变C.313del(P.Ala105fs)和C.199del(p。is67fs)(图) 1 e)。 图1 D)。单核苷酸多态性分析表明,与患者的PBMC相比,BIHI292-A HIPSC线没有任何拷贝数变量> 2 Mb> 2 MB> 5 MB(表1)。短串联重复(STR)分析的结果表明,BIHI292-A细胞系和患者的PBMC的遗传认同是相同的(表1)。Sanger测序证实了两个Exon 2中的两个Trem2杂合突变C.313del(P.Ala105fs)和C.199del(p。is67fs)(图1 e)。图为了确认患者突变的存在(Buthut等,2023),在TREM2基因的外显子2中为复合杂合突变进行了BIHI292-A HIPSC的测序。通过将分化为三个细菌层的细胞进行分化,测试了多能分化势。分化测试证实,BIHI292-A HIPSC具有分化为内胚层(CD184 +,SOX17 +),Meso Dermal(CD140B +,CD144 +)和外胚层(PAX-6 +,SOX2 +)细胞的潜力(1 f)。BIHI292-A HIPSC对支原体进行了阴性测试(Suppl。1 b)。

对“小胶质激活状态驱动神经退行性疾病的葡萄糖摄取和FDG-PET改变”的评论的回应
引起了极大的兴趣,我们认识到Zimmer等人的技术评论。Xiang等人在我们最近的科学转化医学论文中。 (1)。 我们感谢关于氟脱氧葡萄糖正电子发射断层扫描(FDG-PET)信号的细胞起源的讨论,并且考虑到剩下的许多开放问题,我们希望这将刺激该领域的进一步重要研究。 我们完全同意Zimmer等人。 星形胶质细胞在大脑的FDG摄取中起着重要作用,并且我们没有否认星形胶质细胞FDG吸收的贡献是整体FDG-PET信号的主要来源。 但是,考虑到另一组(2)的最新数据,以及我们的发现(1),小胶质细胞FDG摄取对FDG-PET信号的实质性直接贡献,特别是在髓样细胞2(trem2)表达的Trig Gering受体2(trem2) - 依赖性激活的激活非常重要的情况下,重要的是要考虑。 首先,在技术评论中,Zimmer等人。 表明,总小胶质细胞数量可能太低,无法影响总FDG-PET信号。 但是,我们的数据清楚地表明,小胶质细胞是淀粉样小鼠模型中FDG-PET的增加,并且在TREM2-KNOCKOUT(TREM2-KO)小鼠中降低了小胶质细胞,而小胶质细胞占健康小鼠脑中FDG总吸收的约10%。 小胶质细胞对FDG摄取的贡献是由野生型小鼠的99%小胶质细胞耗竭后的PET信号降低的幅度确定的。 我们在分离细胞中绝对活性的交叉校准后重新评估了我们的数据,并在分离前整个大脑的活性。 1,a和b)(3)。 1C)。Xiang等人在我们最近的科学转化医学论文中。(1)。我们感谢关于氟脱氧葡萄糖正电子发射断层扫描(FDG-PET)信号的细胞起源的讨论,并且考虑到剩下的许多开放问题,我们希望这将刺激该领域的进一步重要研究。我们完全同意Zimmer等人。星形胶质细胞在大脑的FDG摄取中起着重要作用,并且我们没有否认星形胶质细胞FDG吸收的贡献是整体FDG-PET信号的主要来源。但是,考虑到另一组(2)的最新数据,以及我们的发现(1),小胶质细胞FDG摄取对FDG-PET信号的实质性直接贡献,特别是在髓样细胞2(trem2)表达的Trig Gering受体2(trem2) - 依赖性激活的激活非常重要的情况下,重要的是要考虑。首先,在技术评论中,Zimmer等人。表明,总小胶质细胞数量可能太低,无法影响总FDG-PET信号。但是,我们的数据清楚地表明,小胶质细胞是淀粉样小鼠模型中FDG-PET的增加,并且在TREM2-KNOCKOUT(TREM2-KO)小鼠中降低了小胶质细胞,而小胶质细胞占健康小鼠脑中FDG总吸收的约10%。小胶质细胞对FDG摄取的贡献是由野生型小鼠的99%小胶质细胞耗竭后的PET信号降低的幅度确定的。我们在分离细胞中绝对活性的交叉校准后重新评估了我们的数据,并在分离前整个大脑的活性。1,a和b)(3)。1C)。我们现在提供了一个额外的FDG分配模型,该模型考虑了示踪剂注射后通过磁性激活的细胞分选评估的单胶质细胞摄取(1),考虑到小鼠大脑中的7%小胶质细胞(图在这里,我们发现了对小胶质细胞对PET(99%耗竭时降低9.8%)和磁性细胞分选的相对贡献的相对贡献(MAC; 8.6%;图。还观察到淀粉样小鼠模型的小胶质细胞对PET(29.1%)和MAC(29.1%)和MAC(29.9%)的总FDG贡献之间的良好一致性,假设由于繁殖而导致的小胶质细胞密度增加了1.3倍(1)。

第二届 DZG 慕尼黑日
10. 夜间高温暴露与中风风险 | 何成(DZHK) 11. 增强的legumain活性将前颗粒蛋白缺乏与额颞叶变性的TDP-43病理联系起来 | Maria-Teresa Mühlhofer(DZNE) 12. 由LMP1-TRAF6复合物直接介导的Epstein-Barr病毒驱动的B细胞淋巴瘤 | Fabian Giehler(DZIF) 13. 小鼠和人脑中前脉冲抑制和精神分裂症基因的共表达 | Lillian Garrett(DZPG) 14. TREM2:从靶标识别到翻译 | Kai Schlepckow(DZNE) 15. 免疫介导的松果体失神经支配是心脏病睡眠障碍的基础 | Karin Ziegler(DZHK) 16. 解剖肌肉中基于Treg的免疫代谢串扰 | Maike Becker (DZD) 17. 肝脏免疫变阻器调节慢性 HBV 感染中的 CD8 T 细胞免疫 | Hannah Wintersteller (DZIF)
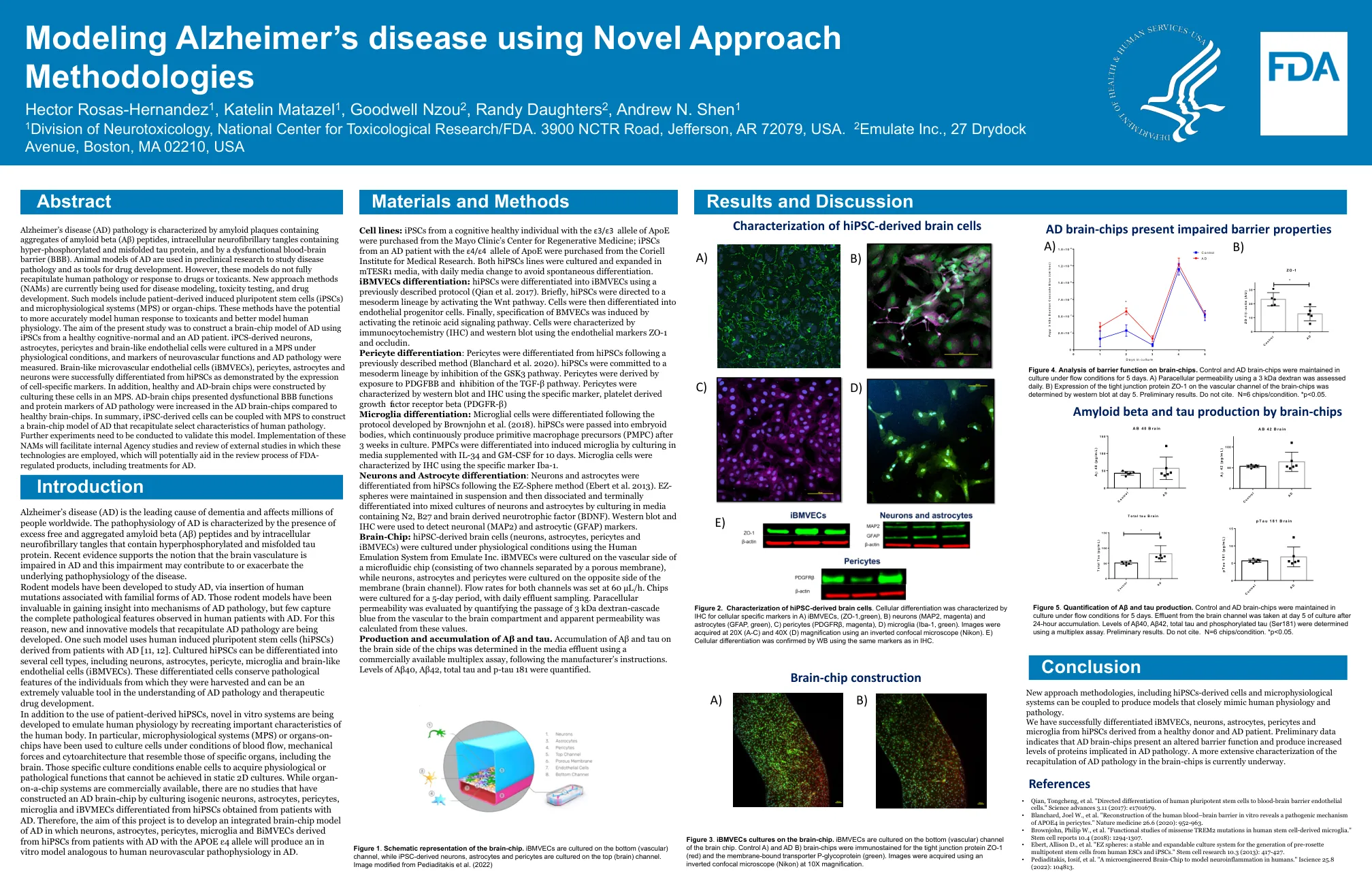
使用新方法方法
i p系统是由brain脑chi iSogeni I小胶质细胞和IBVMEC构建的,与从CE s患者获得的HIPSC区分开来。“体外人类血脑屏障的重建揭示了一种致病机制。因此,该项目的目的是开发周细胞中APOE4的综合脑芯片模型。”自然医学26.6(2020):952-963。神经元,星形胶质细胞,周细胞,小胶质细胞和BIMVEC的AD•BrownJohn,Philip W.等。“人类干细胞衍生的小胶质细胞中错义Trem2突变的功能研究。”来自APOEε4等位基因AD患者的HIPSC的将在图3中产生A。 ibmvecs培养在脑芯片上。 IBMVEC在底部(血管)通道干细胞报告10.4(2018):1294-1307上培养。将在图3中产生A。ibmvecs培养在脑芯片上。IBMVEC在底部(血管)通道干细胞报告10.4(2018):1294-1307上培养。

神经退行性疾病进入功能基因组学时代
低成本 DNA 测序的普及使另一种将人类遗传学与神经系统疾病的特定基因驱动因素联系起来的方法——全基因组关联研究 (GWAS) 复活。通过比较患病个体与未患病(“对照”)人群的基因组成,可以确定增加患病可能性的风险因素。GWAS 导致发现特定基因的变异,包括 TREM2(髓系细胞 2 上表达的触发受体)和 GBA1(葡萄糖脑苷脂酶 1),分别是非孟德尔 AD 和 PD 的风险因素。在某些情况下,GWAS 结果突出了以前被低估的导致发病的机制。例如,与 AD 风险相关的遗传变异在髓系细胞(可能是小胶质细胞)中起作用的基因和增强子(基因组中控制基因表达的区域)中富集。这表明先天免疫细胞在 AD 中发挥着重要作用。因此,特定生物途径中风险变异的丰富可以加深我们对神经退行性疾病的机制理解,甚至可能指出新的治疗目标。

健康与疾病中的小胶质细胞
• Fabp5 mediates lipid metabolism to drive microglia phenotype transition and neuroinflammation following stroke Yan Li • Senescent microglia conserved in aging and Alzheimer's disease exhibit elevated TREM2 protein levels Noa Rachmian • SorLA limits inflammatory properties of microglia during glioma progression Paulina Kaminska • Galectin-3 role in the小胶质细胞和淀粉样蛋白β的相互作用•小胶质细胞的多色命运图显示了缺血性中风后克隆膨胀,异质性和细胞 - 细胞相互作用。Majed Kikhia•纤维淀粉样蛋白-Beta诱导细胞应激,并改变小胶质细胞中的蛋白质降解Alison Carlisle•一种体外平台,通过与毒素性雌雄同体的血管构成血管化的脑巨噬细胞来产生人脑巨噬细胞。Amin Yarmand•神经胶质瘤中的小胶质细胞动力学:一种鉴定与疾病相关的小胶质细胞Jiawen Qian的命运映射方法•通过CSF1R操纵单核吞噬系统,以了解内源机制和增强后创伤和增强。

Syk 抑制剂可防止培养中小胶质细胞介导的神经元丢失
小胶质细胞是脑巨噬细胞,由于其炎症和吞噬活性,在许多脑病理中发挥有益和/或有害作用。小胶质细胞炎症和吞噬作用被认为受脾脏酪氨酸激酶 (Syk) 调节,该酶由多种小胶质细胞受体激活,包括与神经退行性有关的 TREM2(髓系细胞上表达的触发受体 2)。在这里,我们测试了 Syk 抑制剂是否可以预防原代神经元-胶质细胞培养物中脂多糖 (LPS) 诱导的小胶质细胞依赖性神经退行性。我们发现 Syk 抑制剂 BAY61-3606 和 P505-15(分别为 1 µ M 和 10 µ M)完全阻止了 LPS 诱导的神经元丢失,这是小胶质细胞依赖性的。Syk 抑制还可以防止较老的神经元-胶质细胞培养物中神经元的自发丢失。在没有 LPS 的情况下,抑制 Syk 会耗尽培养物中的小胶质细胞并导致一些小胶质细胞死亡。然而,在存在 LPS 的情况下,抑制 Syk 对小胶质细胞密度的影响相对较小(减少 0-30%),而对两种促炎细胞因子的释放有相反的影响(IL-6 减少约 45%,TNF α 增加 80%)。抑制 Syk 对暴露于 LPS 的小胶质细胞的形态转变也没有影响。另一方面,抑制 Syk 会降低小胶质细胞对珠子、突触和神经元的吞噬作用。因此,在该模型中抑制 Syk 很可能通过减少小胶质细胞吞噬作用来发挥神经保护作用,然而,小胶质细胞密度降低和 IL-6 释放也可能有一定作用。这项研究进一步证明,Syk 是小胶质细胞对神经退行性疾病贡献的关键调节器,并表明 Syk 抑制剂可用于防止小胶质细胞过度吞噬突触和神经元。

下载
额颞痴呆是一种异质性神经退行性疾病,其特征是额叶和颞叶中的神经元丧失。尽管在理解哪些基因与额颞痴呆的病因相关的过程中取得了进展,但这些基因突变如何导致特定皮质区域的细胞损失的生物学基础尚不清楚。在这项工作中,我们合并了来自艾伦脑科学研究所的16个772基因的基因表达数据,并在遗传型额叶痴呆型起始研究中获得了Symp-Tomatic C9orf72,GRN和MAPT突变载体中灰质萎缩的脑图。在C9orf72,GRN和MAPT表达之间以及各个遗传群中的萎缩模式之间没有看到任何signi-ni-firstossion。调整了空间自相关后,1000至5000个基因在每个遗传组中的萎缩模式存在负或正相关,其中最显着相关的基因为TREM2,SSBP3和GPR158(C9orf72,GRN和MAPT和MAPT和MAPT中的MAPT,MXRA和LPA(C9ORF)和LPA(C9orf72中的负相关)(分别为c9orf72),mxra8和lpa(C9OR)(C9) 分别)。过分代表分析确定了与线粒体功能所涉及的基因的负相关性,以及与每个遗传组中血管和神经胶质细胞功能的基因的阳性关联。一组423和700个基因在所有三个地图中均与萎缩模式分别显示出显着的阳性和阴性关联。在富含皮质区域的表达增加的基因富含神经元和小胶质细胞基因,而在萎缩区域中表达增加的基因富含星形胶质细胞和内皮细胞基因。我们的分析表明,这些细胞类型在额颞痴呆中的神经退行性发作中可能比以前假定的神经退行性发作更为活跃,并且对于正相关的细胞标记基因,可能通过神经毒性星形胶质细胞的出现和血液 - 脑障碍物的变化而有可能出现。

解锁罕见疾病遗传学:从基因组中的见解 -
解锁罕见疾病遗传学:全基因组关联研究和单核苷酸多态性的见解Osama Alam* 1,卫生汗2,Azmat Ullah 1 1 1 1 1 1科学与技术大学生物技术系Bannu Bannu,28100 Khyber Pakhtunkhwa,Pakhtunkhwa,Pakhthwa,pakistan。2 govt动物学系。巴基斯坦班努大学研究生学院。摘要:全基因组关联研究(GWAS)是识别与复杂疾病相关的遗传变异的强大工具。然而,由于这些疾病通常涉及低频基因突变,因此它们的效用在阐明稀有疾病的遗传学方面受到限制。代表基因组中单碱基对变化的单核苷酸多态性(SNP)可以为稀有疾病的遗传结构提供宝贵的见解。值得注意的是,诸如APP,PSEN1,PSEN2,APOE,TREM2和ABCA7之类的基因中的特定SNP揭示了阿尔茨海默氏病(AD)的分子基础。App Gene中的SNP RS429358中,通过改变淀粉样蛋白β的产生,与阿尔茨海默氏症的风险增加相关。同样,通过GWAS发现的SNP与慢性阻塞性肺疾病易感性(COPD),纤维肿瘤肿瘤易感性(FOP)和Hutchinson-Gilford综合征(HGPS)联系起来,也表现出ACVR1和LMNA中引起疾病的突变。然而,由于招募大型队列的挑战,GWAS进行了有力的罕见疾病,这给人带来了困难。标准GWAS工作流程涉及患者入学,基因组DNA提取,基因分型和严格的质量控制。案例和对照进行匹配和分析,并进行多次测试的校正。罕见的变体方法和插入旨在增强统计能力的目的。关键障碍包括样本量不足,遗传异质性和罕见的因果变体。缓解策略结合了跨国联盟,基于家庭的设计,功能分析,下一代测序,定制基因面板和机器学习方法。前进的GWA将需要越来越大,多样化的数据集,以及新颖的统计和高通量的OMIC技术,以破译稀有和复杂病理的遗传根。