XiaoMi-AI文件搜索系统
World File Search SystemWildType

MDM2 抑制联合内分泌治疗和 CDK4/6 抑制治疗 ER 阳性乳腺癌
背景:内分泌治疗耐药是雌激素受体 (ER) 阳性乳腺癌治疗的主要临床挑战。在这种情况下,p53 通常是野生型,其活性可能通过上调其关键调节因子 MDM2 而受到抑制。这构成了我们评估 MDM2 抑制作为治疗耐药性 ER 阳性乳腺癌的治疗策略的理论基础。方法:我们使用 MDM2 抑制剂 NVP-CGM097 单独治疗体外和体内模型以及与氟维司群或哌柏西利联合治疗。我们进行细胞活力、细胞周期、凋亡和衰老测定,以评估 p53 野生型和 p53 突变 ER 阳性细胞系 (MCF-7、ZR75-1、T-47D) 以及对内分泌治疗和 CDK4/6 抑制有耐药性的 MCF-7 系中的抗肿瘤作用。我们进一步评估了药物在内分泌敏感和内分泌抗性的 ER 阳性乳腺癌患者异种移植 (PDX) 模型中的作用。

乳酸受体 HCAR1 调节新生儿缺氧缺血后的神经发生和小胶质细胞活化
摘要 新生儿脑缺氧缺血 (HI) 是新生儿死亡和残疾的主要原因,目前唯一的治疗方法是低温疗法。深入了解促进 HI 后组织修复的途径可能有助于开发更好的治疗方法。在这里,我们研究了乳酸受体 HCAR1 在小鼠新生儿 HI 后组织修复中的作用。我们发现与野生型小鼠相比,HCAR1 基因敲除小鼠的组织再生减少。此外,神经祖细胞和神经胶质细胞的增殖以及小胶质细胞活化受损。转录组分析显示,野生型小鼠脑室下区对 HI 的转录反应强烈,涉及约 7300 个基因。相比之下,HCAR1 基因敲除小鼠表现出适度的反应,涉及约 750 个基因。值得注意的是,在 HCAR1 基因敲除中,组织修复的基本过程(如细胞周期和先天免疫)失调。我们的数据表明 HCAR1 是促进 HI 后组织再生的途径的关键转录调节因子。

gαs对于β-arrestin耦合是可拨的,但决定了GRK ...
β -arrestin在G蛋白 - 耦合受体(GPCR)内在化,传统和信号传导中起关键作用。β-抑制蛋白是否独立于G蛋白 - 介导的信号传导尚未完全阐明。使用基因组编辑的研究的研究表明,G蛋白对于通过GPCRS的促丝分裂原激活蛋白激酶激活至关重要,而β-抑制蛋白在信号分区 - 室化中起更为重要的作用。然而,在没有G蛋白的情况下,GPCR可能不会激活β -arrestin,从而限制了将G蛋白与β -arrestin介导的信号事件区分开的能力。我们使用β2-肾上腺素能受体(β2AR)及其在人类胚胎肾脏中表达的β2AR-C尾突变体293个细胞野生型或CRISPR - CAS9基因 - cas9基因编辑,编辑为GαS,β-arrestin1/2,或GPCR ki-Nases 2/3/5/6组合的群体结合量的cas9基因 - 控制基因表达中的暂停。我们发现,β2AR和β-甲素构象变化,β-甲素的募集和受体内在化不需要GαS,但是GαS决定了参与β-arrestin募集的GPCR激酶。通过RNA-Seq分析,我们发现蛋白激酶A和有丝分裂原活化的蛋白激酶基因信号通过刺激野生型和β2AR在野生型和β-arrestin1/2-kO细胞中激活,但在GαS-KO细胞中不存在。 这些结果通过在相应的KO细胞中表达gαs并在野生型细胞中沉降β-阻滞蛋白来验证。 这些发现扩展到表达内源性β2AR水平的细胞系统。通过RNA-Seq分析,我们发现蛋白激酶A和有丝分裂原活化的蛋白激酶基因信号通过刺激野生型和β2AR在野生型和β-arrestin1/2-kO细胞中激活,但在GαS-KO细胞中不存在。这些结果通过在相应的KO细胞中表达gαs并在野生型细胞中沉降β-阻滞蛋白来验证。这些发现扩展到表达内源性β2AR水平的细胞系统。总体而言,我们的结果支持GS对于β2AR促进的蛋白激酶A和有丝分裂原激活的蛋白激酶基因表达特征至关重要,而β-arrestins启动了调节GαSS驱动核转录活性的信号传导事件。

PBFT02 的非临床和早期临床开发,这是一种针对 GRN 突变 FTD 的 AAV 基因疗法(FTD- GRN)
## p < 0.01;### p < 0.005 vs WT;* p < 0.05,*** p < 0.005 vs Grn –/– + V,单因素方差分析,然后进行 Tukey 多重比较检验。缩写;GRN,颗粒蛋白基因;ICV,脑室内;V,载体;WT,野生型 Grn –/– 和 WT 小鼠(n=14-15/gp)ICV 给药 PBFT02 或载体(V)。基线对照是第 1 天未经治疗的小鼠。条形图:平均值 +/- SEM。
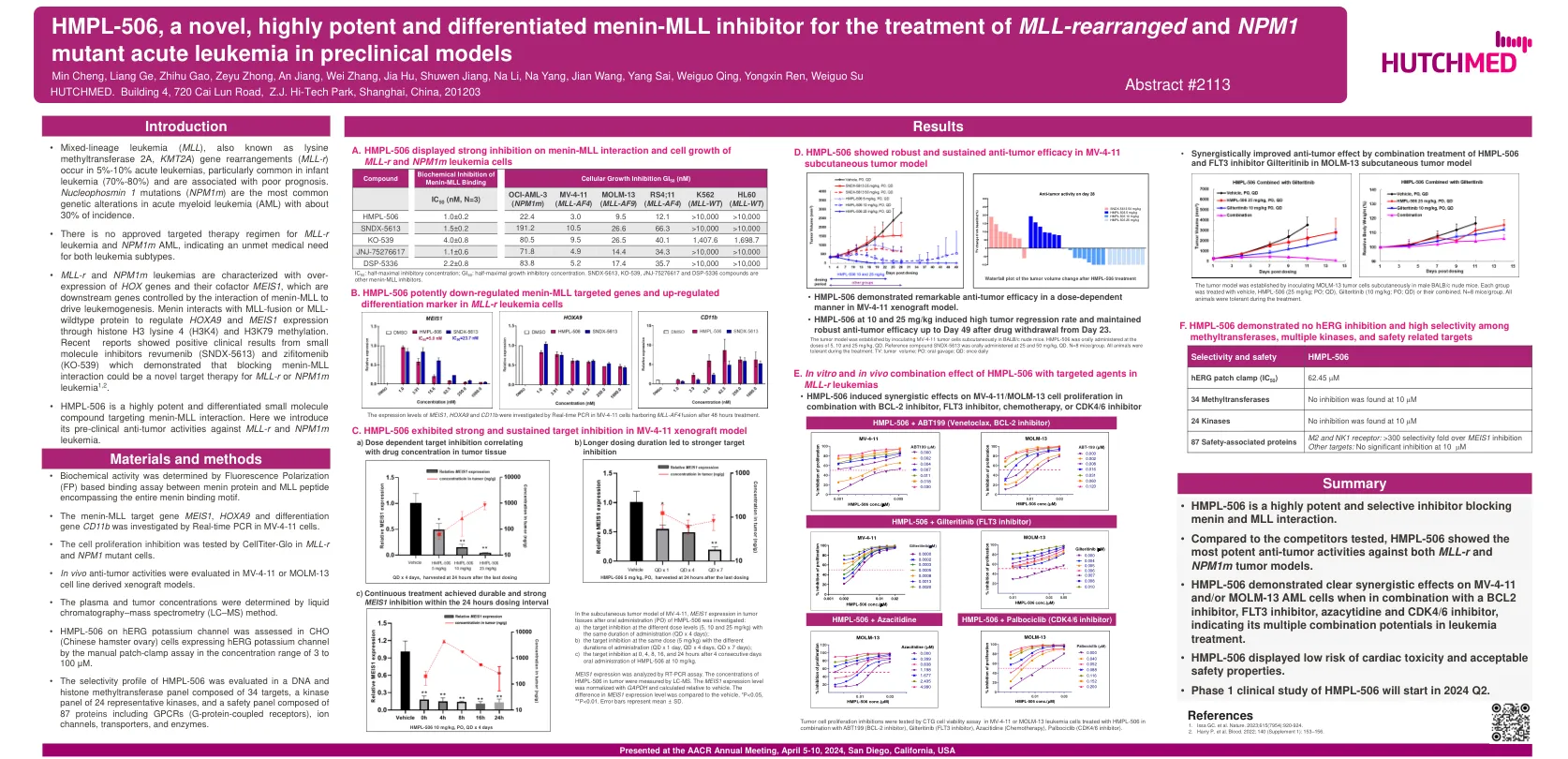
HMPL-506,一部小说,高度有效的和分化的Menin ...
HOX基因及其辅助因子Meis1的表达,它们是由Menin-Mll相互作用以驱动白血病发生的下游基因。Menin与MLL融合或MLL-野生型蛋白相互作用,通过组蛋白H3赖氨酸4(H3K4)和H3K79甲基化来调节HOXA9和MEIS1表达。最近的报道显示,小分子抑制剂rebumenib(SNDX-5613)和Zifitomenib(KO-539)的阳性临床结果,这些结果表明,阻断Menin-MLL相互作用可能是对MLL-R-R或NPM1M1M1M1M1M1M1M1M1M1,2的新型靶标治疗。

资源中的自然设计的植物微生物组 -
间皮瘤是一种罕见的癌症,运输和采矿人群的死亡率较高。这些患者几乎没有治疗选择,这可能部分归因于肿瘤的化学疗法反应有限。我们最初假设奎诺克氨酸可以与顺铂合并,也可以在协同上消除间皮瘤细胞。与顺铂的组合导致了协同的细胞死亡,尽管新型的人工产生的Pemetrexed耐药细胞对Quinarparion更敏感,但与Pemetrexed的结合并不是协同的。出乎意料的是,我们发现具有NF2突变的细胞对喹分非常敏感。分别通过NF2异位表达和NF2突变体和野生型细胞系中的NF2异位表达和敲低来确定喹分河敏感性的这种变化。在间皮瘤中很少有共同的突变,并且这些肿瘤中有60%存在于60%。我们发现喹啉改变了NF2突变的细胞中3000多个基因的表达,这些基因与NF2野生型细胞的变化明显不同。在mRNA和蛋白质水平上验证了NF2/HIPPO途径生物标志物的变化。此外,喹乳可在NF2突变的细胞中诱导G1相细胞周期停滞,而NF2-WildType细胞中的S相阻滞。这项研究表明,奎诺克林可能对大部分间皮瘤患者具有重新利用的潜力。

卫生技术简报2024年4月
vutrisiran是一种药物,正在开发用于与心肌病(ATTR-CM)的甲状腺素蛋白淀粉样变性的成年人。attr-cm被分类为具有心肌病(WTATTR-CM)的蛋白型跨甲状腺素蛋白(WTATTR)淀粉样变性,或伴有心肌病(HATTR-CM)的遗传性经胸蛋白(HATTR)淀粉样蛋白(HATTR)。attr-cm发生时,当一种称为经甲状腺素蛋白(TTR)的蛋白质变为缺陷和聚集体时,会在心脏和其他器官中形成淀粉样蛋白沉积物。淀粉样蛋白沉积物会损害器官的结构和功能,从而导致心脏组织的增厚和僵硬(心肌病),这会导致严重的疾病并致命。WTATTR形式是由于正常功能的TTR蛋白因衰老而有缺陷而导致的。相比之下,HATTR形式发生是由于家庭中运行的错误基因,并且可以从亲生父母传给其子女。attr-cm是一种良好认可的状况,诊断延迟和预后不良,治疗选择仅限于最佳支持性护理来治疗症状。vutrisiran是一种“小干扰RNA”(siRNA)。这是一块非常短的合成遗传物质,旨在连接和阻断负责产生经甲状腺素蛋白的细胞的遗传物质。这减少了甲状腺素有缺陷的产生,从而减少了淀粉样蛋白和缓解症状的形成。vutrisiran每三个月通过皮肤下的注射来给药。如果获得许可,则Vutrisiran将为遗传或野生型Attr-CM的成年人提供治疗选择。

多能性景观的无偏分析揭示了克隆命运偏差的空间调节因素
尽管 Ptch1 编辑导致祖细胞重新偏向或由于 Hedgehog 通路的功能获得而导致谱系进展严重中断,但针对其他受体可能会导致以更微妙的方式调整克隆组成。我们专注于躯干神经嵴,它一直被热议为是受限制祖细胞的混合群体,还是高度多能干细胞的群体 [39-42]。野生型胚胎的克隆变异分析显示 375

