XiaoMi-AI文件搜索系统
World File Search SystemWildType

使用 Cas12a 核酸酶的下一代 CRISPR 基因驱动系统
图 1. 基于 Cas12a 的基因驱动显示出受温度调节的超孟德尔遗传率。(a)CopyCat 基因驱动系统示意图。DsRed 标记的 Cas12a 是一种静态转基因,它通过等位基因转换提供复制 GFP 标记的 CopyCat 元素的核酸酶,而等位基因转换由周围的同源臂驱动。(b)表达 Cas12a 的雄性与携带黑檀木 CopyCat 构建体(e1 或 e4 基因驱动)的处女雌性杂交方案。收集的处女雌性(Cas12a-dsRed + 基因驱动-GFP)与黑檀木突变雄性杂交,通过筛选 F2 后代中的 GFP 标记来评估种系传递率。深灰色半箭头表示雄性 Y 染色体。F1 雌性中的绿色三角形表示潜在的基因驱动复制到野生型染色体上。 (c) 通过对 GFP 标记的乌木 CopyCat 构建体的 F2 后代进行表型评分,评估 F1 雌性生殖系中的基因驱动活性。遗传率测量值与平均遗传率 (%)(也以黑条表示)和进行的 F1 杂交次数 (n) 一起报告在图表顶部。

RSPO3介导的代谢肝分离化调节小鼠的全身葡萄糖代谢和体重
非整倍性通常对细胞存活和生长构成挑战。然而,最近的研究发现了异倍性对某些调节基因突变的细胞有益的例外。我们的研究表明,缺乏纺锤体检查点基因BUB3的细胞表现出精选染色体的非整倍性。与野生型细胞相比,BUB3和BUB1的主轴检查点并不是萌芽的酵母,但BUB3和BUB1的损失增加了Chro Mosome错误分析的可能性。与普遍的假设相反,即由于生长缺陷,非整倍性细胞将胜任,我们的发现表明,bub3δ细胞在许多世代中始终保持特定染色体的脑倍倍倍。我们研究了这些额外的Chromo躯体在BUB3δ细胞中的持久性是由某些基因的有益表达升高而导致的,还是仅仅是耐受性。我们确定了涉及染色体分离和细胞周期调节的几个基因,这些基因赋予了对Bub3缺乏细胞的优势。总的来说,我们的结果表明,特定基因通过非整倍性的增益可能为染色体隔离保真度较差的菌株提供生存优势。

通过结合机器学习和超高通量筛选,工程高度活跃和多样化的核酸酶酶
设计酶以在新型化学环境中起作用是合成生物学具有广泛应用的核心目标。在这项工作中,我们描述了一项由机器学习(ML)引导的运动,以设计核酸酶NucB,核酸核酸核酸hut(一种酶)在治疗慢性伤口时应用。在多轮酶演化运动中,我们将超高通量功能筛选与ML相结合,并将其与维特罗定向进化(DE)的平行运动(DE)和硅内命中率重组(HR)进行了比较。ML引导的运动发现了数百种高度活跃的变体,最多有19倍的核酸酶活性改善,表现优于DE发现的12倍改进。此外,ML设计的命中率距离NUCB WildType高达15个突变,在命中率和多样性方面远远超过了HR方法。我们还表明,仅在进化数据上训练的模型而无需访问任何实验数据,就可以比传统的初始图书馆生成方法以明显高的速率设计功能变体。为了推动ML引导设计的未来进展,我们策划了一个55K多种变体的数据集,这是迄今为止最广泛的基因型 - 表型酶活性景观之一。数据和代码可在以下网址提供:https://github.com/google-deepmind/nuclease_design。

通过结合机器学习和超高通量筛选,工程高度活跃和多样化的核酸酶酶
设计酶以在新型化学环境中起作用是合成生物学具有广泛应用的核心目标。使用机器学习(ML)引导蛋白质设计有可能通过精确导航坚固的健身景观来加速发现高性能酶。在这项工作中,我们描述了ML引导的运动,以设计Nuclease NucB,该核定是一种酶,该酶在治疗慢性伤口的酶降解生物膜,以治疗慢性伤口。在多发酶演化活动中,我们将超高通量功能筛选与ML相结合,并将其与平行的电脑内定向进化(DE)和硅内命中重组(HR)策略进行了比较。ML引导的运动发现了数百种高度活跃的变体,最多有19倍的核酸酶活性改善,而DE的最佳变体提高了12倍。此外,ML设计的命中率距离NUCB WildType高达15个突变,在命中率和多样性方面远远超过了HR方法。我们还表明,仅在进化数据上训练的模型而无需访问任何实验数据,就可以比传统的初始图书馆生成方法以明显高的速率设计功能变体。为了推动ML引导设计的未来进展,我们策划了一个55K多种变体的数据集,这是迄今为止最广泛的基因型 - 表型酶活性景观之一。数据和代码可在以下网址提供:https://github.com/google-deepmind/nuclease_design。

年度报告2024
生物医学研究所2025-01-06奖学金宣布 - 博士后同学项目标题:泛素蛋白 - 依比Quitasin sigase huwe1在免疫项目持续时间和日期中的作用和机制:6个月,01.03.2025-31.08.08.2025应用程序申请书: Anetta.hartlova@gu.se项目摘要:背景天生免疫系统要求严格的法规以确保对病原体的有效防御而不会引起自我伤害。泛素化是一种关键的翻译后修饰,它控制了包括先天免疫信号通路在内的差异细胞过程。泛素化涉及通过E3泛素连接酶将泛素肽偶联到靶蛋白上。尚未完全了解参与先天免疫调节的E3泛素连接酶。最近,我们确定E3连接酶Huwe1是先天免疫的基本调节剂,并且这种酶在小鼠中的消融导致保护与年龄相关的炎症。目的该项目的目的是研究E3连接酶在调节不同先天免疫途径及其对年龄相关炎症和生理功能下降的影响中的作用和机制。方法我们将区分骨髓巨噬细胞和小鼠胚胎成纤维细胞与WildType和Huwe1敲除小鼠,并将在应激挑战时进行比较其细胞因子反应,包括特定先天免疫途径的定义配体。申请应通过电子邮件发送至:anetta.hartlova@gu.se申请应包括:
抑制ULK1/2介导的自噬增强抗原加工和表现 ARC-9:一项评估基于伊特鲁马德的治疗组合的随机研究
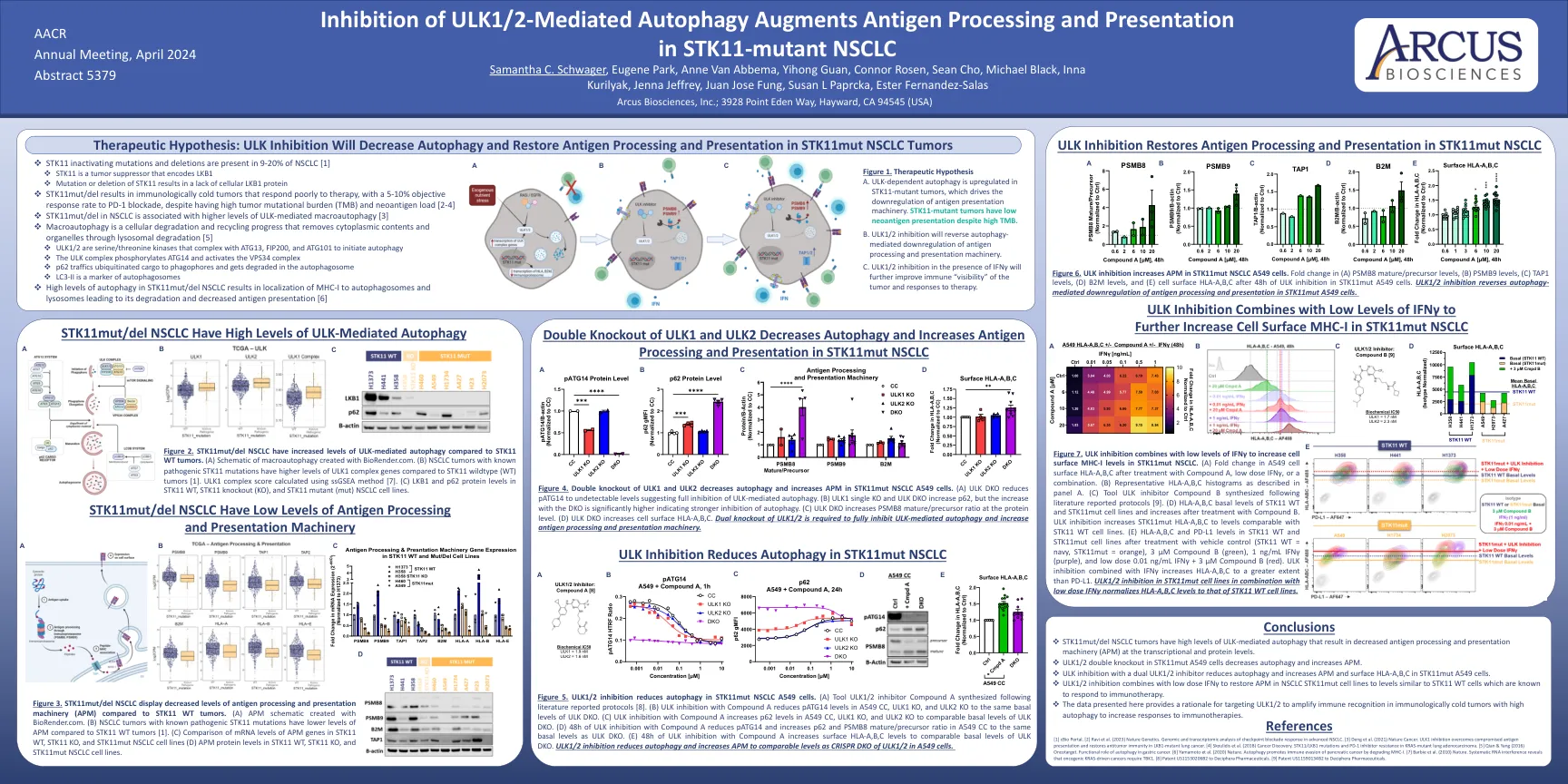
图2。与STK11 WT肿瘤相比, STK11MUT/DEL NSCLC具有ULK介导的自噬水平升高。 (a)用biorender.com创建的大噬细胞的示意图。 与STK11野生型(WT)肿瘤相比,具有已知致病性STK11突变的NSCLC肿瘤具有更高水平的ULK1复合基因[1]。 ULK1复合分数使用SSGSEA方法计算[7]。 (C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。 图4。 ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。 (a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。 (b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。 (c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。 (d)ULK DKO增加了细胞表面HLA-A,b,c。 需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。STK11MUT/DEL NSCLC具有ULK介导的自噬水平升高。(a)用biorender.com创建的大噬细胞的示意图。与STK11野生型(WT)肿瘤相比,具有已知致病性STK11突变的NSCLC肿瘤具有更高水平的ULK1复合基因[1]。ULK1复合分数使用SSGSEA方法计算[7]。 (C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。 图4。 ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。 (a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。 (b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。 (c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。 (d)ULK DKO增加了细胞表面HLA-A,b,c。 需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。ULK1复合分数使用SSGSEA方法计算[7]。(C)STK11 WT,STK11敲除(KO)和STK11突变体(MUT)NSCLC细胞系中的LKB1和P62蛋白水平。图4。ULK1和ULK2的双重敲除可降低自噬并增加STK11MUT NSCLC A549细胞中的APM。(a)ULK DKO将PATG14降低至无法检测的水平,表明对ULK介导的自噬完全抑制。(b)ULK1单个KO和ULK DKO增加了p62,但随着DKO的增加,dKO的增加表明自噬抑制更强。(c)ULK DKO在蛋白质水平上增加了PSMB8成熟/前体比率。(d)ULK DKO增加了细胞表面HLA-A,b,c。需要ULK1/2的双重敲除以完全抑制ULK介导的自噬并增加抗原加工和表现机制。

BGB-16673,BTK DEGRADER,克服了目标...
简介•Bruton酪氨酸激酶(BTK)是BCR信号通路的关键组成部分,其长期激活对于各种B细胞恶性肿瘤中的细胞增殖和存活至关重要。通过共价BTK抑制剂(CBTKI)对BTK的抑制作用,例如Ibrutinib,acalabrutinib和Zanubrutinib,已彻底改变了CLL和其他B细胞恶性肿瘤的管理。•但是,在半胱氨酸481处经常获得BTK抗性突变,该突变消除了CBTKI结合能力,而其他诱导激酶过度活化或激酶独立功能的突变限制了长期临床益处。•非共价BTK抑制剂(例如,折叠曲替尼)在CBTKIS²上进展的BTK C481突变的CLL患者表现出了有希望的功效。即使这样,某些患者也出现了BTK C481以外的BTK突变。•可以解决来自共价BTKI和非共价BTKI的耐药性突变的药物可能会提供新颖的治疗选择。此外,尽管对某些侵袭性淋巴瘤的BTK依赖性有充分的文献证明,但批准的BTKI的临床益处似乎是适中的,并且有必要进一步进行临床研究。具有BTK靶向降解的化合物可能比BTK抑制那些侵略性疾病更具优势。•BGB-16673是一种口服BTK靶向嵌合降解激活(BTK-CDAC)化合物,旨在降解WildType BTK和多突变体形式。目前正在两期研究(NCT05006716,NCT05294731)中进行研究。

非O1/non ...
摘要:非O1和非O139弧菌霍乱(NOVC)会引起人类胃肠道感染。被污染的食物,尤其是海鲜,是人类感染的重要来源。在这项研究中,从零售海鲜中分离出的63个NOVC菌株的毒力潜力在基因型和表型水平上被表征。尽管没有菌株编码霍乱毒素(CTX)和毒素调节的pilus(TCP),但包括Hlya Hymolysin,cholix Toxin CHXA,热稳定的肠毒素STN,以及针对3型和6型分泌系统编码的基因。所有菌株均表现出针对人和绵羊红细胞的溶血活性:90%(n = 57)形成强生生物膜,52%(n = 33)在37℃时高度运动,只有8%(n = 5)和14%(n = 9)可以抗拒60%和≥40%的人类血清。生物膜形成和毒素调节基因。CGMLST分析表明,来自临床NOVC菌株的海鲜簇的NOVC菌株。抗菌易感性测试(AST)导致对五种菌株的鉴定,这些菌株针对β-内酰胺类(包括青霉素,碳碳素,碳酸苯甲酸酯和头孢菌素),多酰氧蛋白,多酰氧蛋白和硫酰胺和硫酰胺的物质产生了非wildtype表型(中和耐药性)。表型抗性模式可以部分归因于在计算机分析中通过鉴定的获得的耐药性决定因素。我们的结果表明,从零售海鲜产品中分离出的分析的NOVC的毒力潜力差异,可以考虑进一步的致病性评估以及对未来海鲜监测中NOVC分离株的风险评估。

自私的隔离变形者超值 - 驱动,重组和遗传负荷
抽象的减数分裂驱动超级基因是链接基因座等位基因的复合物,共同颠覆了孟德尔的隔离,从而产生了优先传播。在男性中,最常见的驱动器机制涉及一对替代等位基因之一的精子的破坏。虽然至少两个基因座对于雄性驱动器(驱动器和目标)很重要,但连接的修饰符可以增强驱动器,从而产生抑制重组的选择压力。在这项工作中,我们研究了常染色体,多焦点,男性减数分裂驱动系统,果蝇果蝇果蝇中的隔离变形(SD)的发展和基因组后果。在非洲人群中,主要的SD染色体变体SD-MAL的特征是两个重叠的,对染色体ARM 2R上的偏心反转,几乎完美(〜100%)传播。我们详细研究了SD-MAL系统,探索其成分,染色体结构和进化史。我们的发现表明,最近的染色体规模的选择性扫描是由强烈的上位型选择的单倍型,主要驾驶等位基因,主要驾驶等位基因和一个或多个因素。尽管大多数SD-MAL染色体都是纯合子致死的,但SD-MAL单倍型可以与其他染色体重组,并通过交叉通过基因转换与Wildtype染色体补充单倍型。SD-MAL染色体具有累积的致命突变,过量的非同义突变和过量的转座元件插入。因此,SD-MAL单倍型作为一种小的半分离亚群演变,具有强烈的选择史。这些结果可以解释世界各地不同人群中SD单倍型的进化周转,并广泛地暗示了超速进化。

NDP 外显子 2 区域 KO 报告模板...
图 S01:CRISPR/Cas9 编辑人类 iPSC。(A) 使用双 sgRNA(sg1 和 sg2)靶向外显子 2 两侧的区域来生成 HDR 介导的 NDP KO-GFP 报告系,使用带有 KO 报告模板的 PUC57 载体,由外显子 2 (5'UTR)-eGFP-P2A 盒组成,两侧是同源臂 (HA),其中包括 PAM 位点突变和用于质粒线性化的 sgRNA 切割位点 (sg1-c 和 sg2-c)。在此过程中还生成了 NHEJ 介导的 NDP KO (NDP Δ exon2) 以及 NDP WT 同源克隆。(B) 使用类似方法生成 NDP WT-GFP 报告系。外显子 3 的 CDS 两侧的双 sgRNA(sg3 和 sg4)和含有修复模板的质粒,由外显子 3(CDS)-V5-P2A- eGFP 盒组成,两侧是同源臂,包括 PAM 位点突变和用于质粒线性化的 sgRNA 切割位点(sg3-c 和 sg4-c)。sgRNA,小向导 RNA;HDR,同源定向修复;KO-GFP,敲除报告基因;KO,敲除;WT,野生型;PAM,protoscpacer 相邻基序;UTR,非翻译区;CDS,编码序列;HA,同源臂;eGFP,增强型绿色荧光蛋白;P2A,猪 teschovirus-1 2A 自切割肽;V5,V5 标签。