XiaoMi-AI文件搜索系统
World File Search Systemarticles

MIT 开放存取文章网络视角...
摘要 当我们学习时,大脑中会发生什么?自从 Cajal 的开创性工作以来,该领域已经取得了许多发现,表明经验如何改变单个突触的结构和功能。然而,最近的进展强调了从神经元和突触群体之间复杂的相互作用来理解学习的必要性。我们应该如何在如此宏观的层面上思考学习?在这里,我们开发了一个概念框架来弥合学习运作的不同尺度之间的差距——从突触到神经元再到行为。利用这个框架,我们探索指导跨这些尺度的感觉运动学习的原则,并为该领域未来的实验和理论工作奠定基础。关键词 神经元群体、感觉运动学习、状态空间框架、神经可塑性、维度、内部模型

MIT 开放存取文章部署策略......
随着太空民主化的兴起,地球观测 (EO) 图像对各行各业来说变得越来越重要。然而,构建能够实现持续高质量全球覆盖的星座仍然困难且成本高昂。将卫星星座重新配置到不同的轨道平面以改变其观测性能传统上是一个燃料密集型过程。可重构星座 (ReCon) 的概念考虑了在进行燃料效率高的机动以改变卫星地面轨道时的 퐽 2 扰动效应。与不可重构星座相比,ReCon 通过减少按需对给定地面事件进行重复观测所需的卫星数量,降低了高重访频率、高质量分辨率、EO 星座的成本。本文首先探讨了 ReCon 性能对重构需求、设计成本和图像价值的不确定性的敏感性。敏感性分析表明,在需求极低的情况下,ReCon 无法提供具有成本效益的解决方案(就每美元花费所响应的事件而言)。在需求高的情况下,ReCon 根本无法满足需求。对一系列需求情景的蒙特卡罗分析表明,使用分阶段部署 ReCon 为应对 EO 图像需求的不确定性提供了一种灵活、具有成本效益的解决方案。通过分阶段部署将发射成本推迟到未来,不仅可以为星座设计提供灵活性,而且还允许设计人员利用持续降低发射成本和增加发射机会的机会。分阶段部署星座还可以使卫星技术随着时间的推移而发展,从而有助于捕获更高价值的图像并进一步增强 ReCon 的功能。实施分阶段部署更多卫星的选项使 ReCon 能够更好地应对太空资产需求的不确定性。

MIT开放访问文章神经科学的神经科学...
瑜伽作为一种实践和生活哲学已有4500多年的历史,并在印度河谷文明中具有瑜伽习俗的已知证据。过去几十年来,瑜伽和冥想的效用是一种实践,其背后越来越多的科学证据。已经发表了重要的科学文献,说明了瑜伽实践的好处,包括“ asana”,“ pranayama”和“ dhyana”对心理和身体健康。电生理和最新功能磁共振成像(fMRI)研究发现了明确的瑜伽实践神经特征。在本文中,我们根据dailistic的“ sankhya'学校进行了对瑜伽哲学的回顾,这适用于帕坦贾利在他的瑜伽经文中摘要的意识,然后对五个“ vritti”(心理模式)的讨论,“ pratyahara”,“ dharaana”,“ dharaana”,“ dhyhara”,“ dhyhara”,“ dhyhara”,“ dyharana”,“ dhyhara”,'''''''''''; ‘samapatti'。我们制定了意识理论(YTC),这是一种具有凝聚力的理论,可以对外部调制和内部状态进行建模。我们提出,应该将注意力,睡眠和思想流浪理解为心灵的独特调节状态。ytc允许我们对外部状态,冥想的内部状态,“ samadhi”甚至意识障碍进行建模。此外,我们列出了一些可检验的神经科学假设,这些假设可以使用YTC回答并分析益处,结果和可能的局限性。

J.拜伦·麦考密克(J.
媒体对有色人种的描述是一个非常重要的问题。尽管占美国人口的近30%,但这些网络要么广播有色人种的刻板印象描述或图像不足。从住房歧视法学借款,本文建议F.C.C.在“普通观众测试”下分析少数族裔不足或刻板印象的案例。普通观众将是合理的人。根据该测试,如果没有特定种族和族裔的代表,广播公司将违反《通信法》。此外,如果有证据表明种族或族裔被描绘成罪犯或恶棍,那么差异也将是违法行为。歧视的意图对于寻找违规行为是不必要的。作为频谱的被许可人,广播公司将其作为所有美国人民的受托人,他们必须以非歧视性的方式使用它。因此,普通的观众测试将确保

MIT开放访问文章工程耦合...
引用:Hart,Stephanie M.等。“使用合成DNA支架进行激子传输的工程耦合。”Chem 7,3(2021年3月):752-773。©2020 Elsevier Inc

文章Melphalan和Presnisone加上沙利度胺与...
自1960年代以来,与Melphalan和泼尼松的结合化疗已被用于多发性骨髓瘤的治疗,并且仍然是不符合高剂量治疗的老年患者的最广泛接受的治疗选择。1,2与烷基化剂的复杂组合通常会增加毒性作用,并增加了不便而没有提供生存优势。3个基于地塞米松的方案也显示出与老年患者的Melphalan和泼尼松方案相比,没有生存优势,并且与毒性更大的作用有关。4,由于与梅尔法兰(Melphalan)和泼尼松的中位生存率约为3年,因此需要新的治疗方法。创新的药物组合或Melphalan的剂量反应效应可用于实现这一目标。沙利度胺在复发或难治性多发性骨髓瘤中显示出大量的抗肿瘤活性。5在新诊断的多发性骨髓瘤患者中,沙利度胺的有用性尚不清楚。与Melphalan和Melphalan和泼尼松结合使用或掺入高剂量治疗中时,沙利度胺的响应率提高,包括完全反应,

关于泌尿科人工智能的前100篇文章
在医学领域,人工智能(AI)迅速变得突出,其对泌尿科的影响是深远的。与传统方法相比,经过验证和优化的AI可导致更快,更个性化,高效和专注的搜索(1)。AI领域继续迅速发展。患者的关系以及患者的结果正在改善(2)。AI有效地处理大量数据的能力,结合向电子患者记录的转变,导致越来越大的“大数据”集。将来,AI将能够分析和检测新颖的诊断和治疗模式(3)。当前的计算机科学进步已经导致了多个高度复杂的非医疗过程的研究和自动优化。如果正确应用,AI模型的开发可以导致对患者相关数据的更有效的处理和分析,以及对泌尿科患者的优化诊断和治疗(4)。

麻省理工学院开放获取文章 量子传感
“量子传感”描述了使用量子系统,量子特性或量子现象的使用来测量物理量。量子传感器的历史示例包括基于超导量子干扰装置和原子蒸气或原子钟的磁力计。最近,量子传感已成为量子科学和技术领域内的一个独特且快速增长的研究分支,其中最常见的平台是旋转量子矩,捕获的离子和通量量子。该领域将在应用物理和其他科学领域提供新的机会,尤其是在高灵敏度和精确度方面。本综述从感兴趣的实验者的角度介绍了量子传感的基本原理,方法和概念。

MIT 开放获取文章 化学中的人工智能...
此作者接受手稿是未经编辑的同行评审手稿的 PDF 文件,已被接受出版但尚未进行编辑或更正。期刊上发表的官方记录版本保持最新,因此可能与此版本不同。使用条款和重用条款:非商业目的的学术研究,请参阅此处了解完整条款。https://www.springer.com/aam-terms-v1
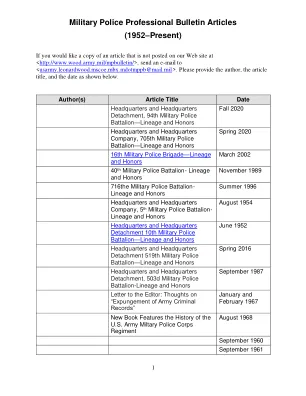
军事警察专业公报文章 (1952 年至今)
如果您想要一份未发布在我们的网站 < http://www.wood.army.mil/mpbulletin/ > 上的文章副本,请发送电子邮件至 < usarmy.leonardwood.mscoe.mbx.mdotmppb@mail.mil >。请提供作者、文章标题和日期,如下所示。