XiaoMi-AI文件搜索系统
World File Search Systembinding

通过绑定界面模拟肽设计
1 中国北京北京大学2号计算机科学技术系2北京北京北京大学AI行业研究所3中国北京7福吉安省脑衰老和神经退行性疾病的关键主要实验室,基础医学科学学院,福建医科大学,福建,富士,富士,纳米材料和纳米安全的纳米材料和纳米安全性生物医学效应的主要实验室中国北京大学 *应致辞:wangzh@iccas.ac.cn,liuyang2011@tsinghua.edu.cn,majianzhu@tsinghua.edu.edu.cn中国北京北京大学2号计算机科学技术系2北京北京北京大学AI行业研究所3中国北京7福吉安省脑衰老和神经退行性疾病的关键主要实验室,基础医学科学学院,福建医科大学,福建,富士,富士,纳米材料和纳米安全的纳米材料和纳米安全性生物医学效应的主要实验室中国北京大学 *应致辞:wangzh@iccas.ac.cn,liuyang2011@tsinghua.edu.cn,majianzhu@tsinghua.edu.edu.cn中国北京北京大学2号计算机科学技术系2北京北京北京大学AI行业研究所3中国北京7福吉安省脑衰老和神经退行性疾病的关键主要实验室,基础医学科学学院,福建医科大学,福建,富士,富士,纳米材料和纳米安全的纳米材料和纳米安全性生物医学效应的主要实验室中国北京大学 *应致辞:wangzh@iccas.ac.cn,liuyang2011@tsinghua.edu.cn,majianzhu@tsinghua.edu.edu.cn中国北京北京大学2号计算机科学技术系2北京北京北京大学AI行业研究所3中国北京7福吉安省脑衰老和神经退行性疾病的关键主要实验室,基础医学科学学院,福建医科大学,福建,富士,富士,纳米材料和纳米安全的纳米材料和纳米安全性生物医学效应的主要实验室中国北京大学 *应致辞:wangzh@iccas.ac.cn,liuyang2011@tsinghua.edu.cn,majianzhu@tsinghua.edu.edu.cn中国北京北京大学2号计算机科学技术系2北京北京北京大学AI行业研究所3中国北京7福吉安省脑衰老和神经退行性疾病的关键主要实验室,基础医学科学学院,福建医科大学,福建,富士,富士,纳米材料和纳米安全的纳米材料和纳米安全性生物医学效应的主要实验室中国北京大学 *应致辞:wangzh@iccas.ac.cn,liuyang2011@tsinghua.edu.cn,majianzhu@tsinghua.edu.edu.cn

辐射感受器的结合曲线,非典型抗精神病药奥氮平
多巴胺D2受体中的神经肌肉处理精神分裂症患者的纹状体(Farde等人1989)。 与典型的抗精神病药不同,氯氮平是一种二苯二氮卓类药物,被发现是一种有效的抗精神病,很少会产生EPSS或迟发性屈服难题(Casey 1989; Claghorn等人; Claghorn等人。 1987),因此被称为非典型。 此外,氯氮平独特地降低了精神分裂症的阴性症状以及阳性症状,并活跃于一部分耐药患者中(Kane等人 1988)。 氯氮平非典型性质的解释重点是与多巴胺D2以外的几种神经元受体的相互作用,包括多巴胺D1(Andersen等人 1986),多巴胺04(van tol等人 1991),血清毒素(SHT)2a(Meltzer等人 1989),SHT2C(Canton等人 1990; Roth等。 1992)和毒蕈碱亚型(Miller and Hiley 1974)。 此外,最近据报道,氯氮平对克隆的SHT 6和SHT7受体具有很高的影响(Roth等人 1994)。 氯氮平是一种有效的抗精神病药,可以被视为非典型抗精神病药的原型,但其使用1989)。与典型的抗精神病药不同,氯氮平是一种二苯二氮卓类药物,被发现是一种有效的抗精神病,很少会产生EPSS或迟发性屈服难题(Casey 1989; Claghorn等人; Claghorn等人。1987),因此被称为非典型。此外,氯氮平独特地降低了精神分裂症的阴性症状以及阳性症状,并活跃于一部分耐药患者中(Kane等人1988)。 氯氮平非典型性质的解释重点是与多巴胺D2以外的几种神经元受体的相互作用,包括多巴胺D1(Andersen等人 1986),多巴胺04(van tol等人 1991),血清毒素(SHT)2a(Meltzer等人 1989),SHT2C(Canton等人 1990; Roth等。 1992)和毒蕈碱亚型(Miller and Hiley 1974)。 此外,最近据报道,氯氮平对克隆的SHT 6和SHT7受体具有很高的影响(Roth等人 1994)。 氯氮平是一种有效的抗精神病药,可以被视为非典型抗精神病药的原型,但其使用1988)。氯氮平非典型性质的解释重点是与多巴胺D2以外的几种神经元受体的相互作用,包括多巴胺D1(Andersen等人1986),多巴胺04(van tol等人1991),血清毒素(SHT)2a(Meltzer等人1989),SHT2C(Canton等人 1990; Roth等。 1992)和毒蕈碱亚型(Miller and Hiley 1974)。 此外,最近据报道,氯氮平对克隆的SHT 6和SHT7受体具有很高的影响(Roth等人 1994)。 氯氮平是一种有效的抗精神病药,可以被视为非典型抗精神病药的原型,但其使用1989),SHT2C(Canton等人1990; Roth等。 1992)和毒蕈碱亚型(Miller and Hiley 1974)。 此外,最近据报道,氯氮平对克隆的SHT 6和SHT7受体具有很高的影响(Roth等人 1994)。 氯氮平是一种有效的抗精神病药,可以被视为非典型抗精神病药的原型,但其使用1990; Roth等。1992)和毒蕈碱亚型(Miller and Hiley 1974)。此外,最近据报道,氯氮平对克隆的SHT 6和SHT7受体具有很高的影响(Roth等人1994)。氯氮平是一种有效的抗精神病药,可以被视为非典型抗精神病药的原型,但其使用

使用 Transformer 进行药物靶标结合亲和力预测
摘要 — 药物发现通常很困难、昂贵且成功率低。药物发现和药物再利用的早期阶段的重要步骤之一是确定药物-靶标相互作用。结合亲和力表示药物-靶标对相互作用的强度。在这方面,已经开发了几种计算方法来预测药物-靶标结合亲和力,并且这些模型的输入表示已被证明在提高准确性方面非常有效。虽然最近的模型比第一个模型更准确地预测结合亲和力,但它们需要靶蛋白的结构。尽管人们对蛋白质结构有浓厚的兴趣,但已知序列和实验确定的结构之间存在巨大差距。因此,找到合适的药物和蛋白质序列表示对于药物-靶标结合亲和力预测至关重要。在本文中,我们的主要目标是评估药物和蛋白质序列表示以改进药物-靶标结合亲和力预测。

“脑健康创新外交:一个典范结合...
1 美国德克萨斯州休斯顿贝勒医学院医学院 2 美国华盛顿特区美国卫生与公众服务部美国老龄化管理局/社区生活管理局 3 美国加利福尼亚州洛杉矶加州大学洛杉矶分校塞梅尔神经科学与人类行为研究所精神病学系 4 美国加利福尼亚州旧金山加州大学旧金山分校医学院记忆与衰老中心 5 美国加利福尼亚州旧金山全球脑健康研究所 6 爱尔兰都柏林都柏林圣三一学院 7 美国俄勒冈州波特兰俄勒冈健康与科学大学医学院 8 美国俄勒冈州波特兰州立大学城市与公共事务学院老龄化研究所 9 阿根廷布宜诺斯艾利斯法瓦洛罗大学 INECO 基金会认知与转化神经科学研究所 (INCYT)


DNA与二氧化硅结合:作用中的合作吸附
请注意,由于它们的高负电荷,我们排除了两个裸露的DNA(U DD <0)之间吸引人的可能性。上面的这三个条件可以在物理上理解如下。由于DNA无法单独与二氧化硅结合,因此结合剂和DNA之间的吸引力(条件2)将确保DNA粘在结合剂上,而复合物(DNA+结合剂)与二氧化硅结合。结合剂必须与二氧化硅结合才能发生(条件1)。但是,如果两种结合剂之间存在吸引力,则在两个结合剂之间形成复合物,而不是DNA结合剂复合物(条件3),它在能量上更有利。这将降低DNA的结合概率与二氧化硅。在这里值得一提的是,在这项工作中为参数扫描所选择的范围由我们较早的作品12,43指导,其中进行了广泛的无偏见和偏见的分子动力学模拟(伞采样模拟),以评估参数。在此,由于系统的复杂性,我们无法评估参数的确切值,因此尝试了参数扫描。在上述所有计算中,我们将结合剂与DNA(rθ)的浓度比为5。

冷冻揭示了NAV1的前所未有的结合位点。 ...
摘要电压门控钠(Na V)通道Na V 1.7由于其参与人类疼痛综合征,已被确定为潜在的新型镇痛靶标。然而,临床上可用的Na V通道阻断药物在9个Na V通道亚型中没有选择性,Na V 1.1 – Na V 1.9。此外,当前已知的Na V 1.7亚型选择性抑制剂(芳基和酰基磺胺)的两个已知类别具有不良特征,可能会限制其发育。到这一点理解Na v 1.7抑制剂的酰基磺酰胺类别的结构 - 活性关系,例如临床开发候选GDC-0310的例证,仅基于芳基磺胺酰胺抑制剂的单个共晶体结构,与电压 - 传感 - 感应 - sensing-sensing-sensing-sensing-domain domain domain 4(vsd4(VSD4)。为了推进针对Na V 1.7通道的抑制剂设计,我们使用低温电子显微镜(Cryo-EM)追求高分辨率结合的Na V 1.7-VSD4结构。在这里,我们报告了GDC-0310通过与芳基磺胺酰胺抑制剂类结合姿势正交结合模式与Na V 1.7-VSD4接合,该模式识别Na V通道中的可预见的未知配体结合位点。这一发现实现了一种新型杂种抑制剂系列的设计,该系列桥接了芳基 - 磺胺酰胺结合口袋,并可以产生具有实质性分化的结构和特性的分子。总体而言,我们的研究强调了使用迭代和高分辨率结构引导的抑制剂设计来追求挑战性药物靶标的冷冻EM方法的力量。这项工作还强调了膜双层在优化靶向VSD4的选择性NA V通道调节器中的重要作用。

使用 AlphaFold 进行可溶性感知蛋白质结合肽设计
摘要:新的蛋白质-蛋白质相互作用(PPI)正在不断被发现,但PPI 与传统靶标相比具有不同的物理化学性质,这使得使用小分子变得困难。肽为靶向 PPI 提供了一种新的方式,但通过计算设计合适的肽序列具有挑战性。最近,AlphaFold 和 RosettaFold 使得从氨基酸序列预测蛋白质结构成为可能,并且精度极高,从而实现了从头蛋白质设计。我们使用 AfDesign 的“结合剂幻觉”协议(一种使用 AlphaFold 的从头蛋白质设计方法)设计了可能以 PPI 为靶蛋白的肽。然而,这些肽的溶解度往往较低。因此,我们使用氨基酸的溶解度指数设计了溶解度损失函数,并开发了可感知溶解度的 AfDesign 结合剂幻觉协议。使用新协议设计的序列中肽的溶解度随着溶解度损失函数的权重的增加而增加;此外,它们还捕捉到了溶解度指数的特征。此外,通过对接结合亲和力评估,新协议序列往往比随机或单残基替换序列具有更高的亲和力。我们的方法表明,可以设计出能够结合PPI界面同时控制溶解度的肽序列。
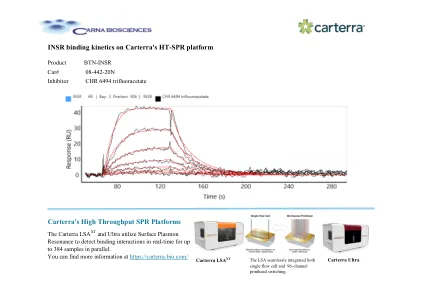
Carterra HT-SPR 平台上的 INSR 结合动力学
Carterra LSA XT 和 Ultra 利用表面等离子体共振实时检测多达 384 个样本的结合相互作用。您可以在 https://carterra-bio.com/ Carterra Ultra 上找到更多信息 LSA 无缝集成了单流动池和 96 通道打印头切换。

基于受体结合域的 COVID-19 疫苗开发
。CC-BY-NC-ND 4.0 国际许可,根据 (未经同行评审认证)提供,是作者/资助者,他已授予 bioRxiv 永久展示预印本的许可。它是此预印本的版权持有者,此版本于 2020 年 5 月 7 日发布。;https://doi.org/10.1101/2020.05.06.079830 doi:bioRxiv 预印本