XiaoMi-AI文件搜索系统
World File Search Systemenhancer

心脏再生需要的损伤响应性MMP14B增强子
哺乳动物的心脏再生能力有限,而斑马鱼具有非凡的再生。在斑马鱼心脏再生期间,内皮细胞可促进心肌细胞周期再入和肌肌的修复,但是负责促进损伤微环境有助于再生的机制仍未完全定义。在这里,我们将基质金属蛋白酶MMP14B识别为心脏再生的主管调节剂。我们确定了斑马鱼和小鼠心脏损伤引起的TEAD依赖性MMP14B内皮增强子,我们表明增强子是再生所必需的,这支持了MMP14B上游的河马信号的作用。最后,我们表明,小鼠中的MMP-14功能对于Agrin的积累很重要,Agrin是新生小鼠心脏再生的基本调节剂。这些发现揭示了促进心脏再生的细胞外基质重塑的机制。

人Rad52刺激RAD51介导的同源搜索
非小细胞肺癌经常在晚期诊断出来,许多患者仍接受经典化学疗法治疗。化学疗法的非选择性通常会导致严重的骨髓抑制。先前的研究表明,蛋白质编码突变无法完全解释骨髓压机的易感性。在这里,我们研究了增强子突变在骨髓抑制易感性中的可能作用。我们生成了三种用卡泊蛋白或吉西他滨处理的三种血管茎的转录组和启动子相互作用图(使用HICAP)。使用公开可用的增强剂数据集的优势,我们使用表观遗传学CRISPR技术验证了硅和活细胞中的HICAP。我们还开发了一种用于相互作用分析和检测差异相互作用基因的网络方法。差异相互作用分析提供了有关相关基因和骨髓抑制途径的其他信息,与散装水平的差异基因表达分析相比。此外,我们表明,与不同水平相关的骨髓抑制水平相关的变体,具有差异相互作用基因的增强子。中心,我们的工作代表了非编码突变的函数注释的整合转录组和基因调节数据集分析的一个突出例子。

心脏再生需要的损伤响应性MMP14B增强子
哺乳动物的心脏再生能力有限,而斑马鱼具有非凡的再生。在斑马鱼心脏再生期间,内皮细胞可促进心肌细胞周期再入和肌肌的修复,但是负责促进损伤微环境有助于再生的机制仍未完全定义。在这里,我们将基质金属蛋白酶MMP14B识别为心脏再生的主管调节剂。我们确定了斑马鱼和小鼠心脏损伤引起的TEAD依赖性MMP14B内皮增强子,我们表明增强子是再生所必需的,这支持了MMP14B上游的河马信号的作用。最后,我们表明,小鼠中的MMP-14功能对于Agrin的积累很重要,Agrin是新生小鼠心脏再生的基本调节剂。这些发现揭示了促进心脏再生的细胞外基质重塑的机制。


CRISPR筛选发现胰腺1
未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。此预印本的版权持有人(该版本发布于4月29日,2024年。; https://doi.org/10.1101/2024.04.26.591412 doi:biorxiv Preprint
Alt-R™ HDR ENHANCER V2 提高效率... - NET
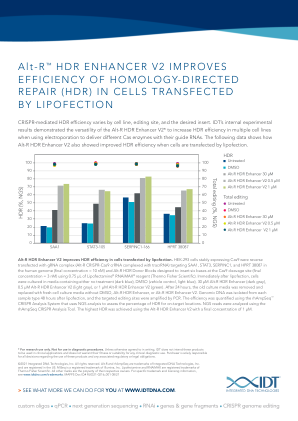
Alt-R HDR Enhancer V2 可提高脂质转染细胞的 HDR 效率。使用 0.75 μL Lipofectamine ® RNAiMAX ® 试剂(赛默飞世尔科技)将稳定表达 Cas9 的 HEK-293 细胞反向转染 gRNA 复合物(Alt-R CRISPR-Cas9 crRNA 与 tracrRNA 复合),靶向人类基因组中的 SAA1、STAT3、SERPINC1 和 HPRT 38087(最终浓度 = 10 nM)和设计用于在 Cas9 裂解位点插入六个碱基的 Alt-R HDR 供体块(最终浓度 = 3 nM)。脂质转染后,立即将细胞培养在含有无处理(深蓝色)、DMSO(载体对照,浅蓝色)、30 μM Alt-R HDR 增强剂(深灰色)、0.5 μM Alt-R HDR 增强剂 V2(浅灰色)或 1 μM Alt-R HDR 增强剂 V2(绿色)的培养基中。24 小时后,移除旧培养基,并用不含 DMSO、Alt-R HDR 增强剂或 Alt-R HDR 增强剂 V2 的新鲜细胞培养基替换。脂质转染 48 小时后从每种样本类型中分离基因组 DNA,并通过 PCR 扩增目标编辑位点。使用 rhAmpSeq ™ CRISPR 分析系统对效率进行量化,该系统使用 NGS 分析来评估目标位置的 HDR 百分比。使用 rhAmpSeq CRISPR 分析工具分析了 NGS 读数。使用最终浓度为 1 μM 的 Alt-R HDR 增强剂 V2 实现了最高的 HDR。

人类加速区域的增强子功能和进化作用
人类加速区域 (HAR) 是人类基因组中进化最快的序列。当 HAR 于 2006 年被发现时,由于非编码基因组的注释很少,它们的功能尚不明了。从转基因动物到机器学习,多种技术一致表明 HAR 可作为基因调控增强子发挥作用,并在神经发育中显著富集。现在可以同时定量测量数千个 HAR 的增强子活性,并模拟每个核苷酸如何促进基因表达。这些策略揭示出许多人类 HAR 序列的功能与黑猩猩直系同源物不同,尽管同一 HAR 中单个核苷酸的变化可能具有相反的效果,与补偿性替换一致。为了全面评估 HAR 在人类进化中的作用,有必要通过实验和计算在更多细胞类型和发育阶段对它们进行剖析。

人类特异性增强子微调径向神经胶质效力和皮质生成
摘要人类进化出一种与发育和基因调节修饰有关的膨胀且复杂的大脑皮层。1-3。人类加速区域(HAR)是具有人类特异性核苷酸取代的高度保守基因组序列。尽管有成千上万的带注释的竖琴,但它们对人类特异性皮质发育的功能贡献在很大程度上是未知的4,5。hare5是在大脑发育过程中活跃的Wnt信号受体Frizzled8(FZD8)的HAR转录增强子6。在这里,使用基因组编辑的小鼠和灵长类动物模型,我们证明了人(HS)Hare5微型皮质发育和连通性通过控制神经祖细胞(NPC)的增殖和神经源能力。HS-HARE5敲入小鼠的新皮质含量显着增大,其中包含更多的神经元。 通过测量体内神经动力学,我们显示了这些解剖学特征与皮质区域之间功能独立性的增加相关。 要了解潜在的发展机制,我们使用实时成像,谱系分析和单细胞RNA测序评估祖细胞命运。 这揭示了HS-HARE5修饰了径向神经胶质祖细胞的行为,在早期发育阶段增加了自我更新,随后神经源性扩大。 我们使用基因组编辑的人和黑猩猩(PT)NPC和皮质器官来评估HS-HARE5和PT-HARE5的相对增强剂活性和功能。 使用这些正交策略,我们显示了HARE5驱动器中的四个人类特异性变体增加了增强剂活性,从而促进了祖细胞增殖。HS-HARE5敲入小鼠的新皮质含量显着增大,其中包含更多的神经元。通过测量体内神经动力学,我们显示了这些解剖学特征与皮质区域之间功能独立性的增加相关。要了解潜在的发展机制,我们使用实时成像,谱系分析和单细胞RNA测序评估祖细胞命运。这揭示了HS-HARE5修饰了径向神经胶质祖细胞的行为,在早期发育阶段增加了自我更新,随后神经源性扩大。我们使用基因组编辑的人和黑猩猩(PT)NPC和皮质器官来评估HS-HARE5和PT-HARE5的相对增强剂活性和功能。使用这些正交策略,我们显示了HARE5驱动器中的四个人类特异性变体增加了增强剂活性,从而促进了祖细胞增殖。这些发现说明了调节性DNA的小变化如何直接影响关键的信号通路和大脑发育。我们的研究揭示了Hars的新功能,这是对人脑皮质的扩张和复杂性至关重要的关键调节元素。

哺乳动物发育增强子激活过程中增强子—启动子相互作用增强
增强子或顺式调控元件可确保在发育过程中对基因表达进行精确的时空控制。该过程由转录因子 (TF) 和辅激活因子介导,它们将调控信息从增强子传递到其目标启动子,跨越的距离可能超过一兆碱基 1-4 。这种增强子-启动子 (E-P) 通讯被认为发生在所谓的拓扑相关结构域 (TAD) 内,拓扑相关结构域是通过黏连蛋白和 CCCTC 结合因子 (CTCF) 的环挤压过程形成的基因组基本组织单位 5-7 。TAD 或 TAD 内染色质相互作用的破坏可能导致基因表达或基因激活的错误下调,并可能导致人类疾病,这表明正确的 E-P 通讯对基因激活的重要性 8-10 。

M01 作为一种专门针对血脑屏障的新型药物增强剂。
对于大多数药物来说,血脑屏障 (BBB) 限制了药物向大脑的输送,而血脑屏障中 claudin-5 决定着内皮旁收缩。为了绕过 BBB,我们将化合物 M01 确定为 claudin-5 相互作用的抑制剂。M01 会导致 BBB 暂时通透,具体取决于不同细胞培养模型中 3 到 48 小时内小分子的浓度。在小鼠中,大脑对荧光素的吸收在注射 M01 后的前 3 小时内达到峰值,并在 48 小时内恢复正常。与单独的细胞抑制紫杉醇相比,M01 改善了紫杉醇向小鼠大脑的输送,并减少了原位胶质母细胞瘤的生长。M01 与 claudin-5 相互作用的结果被纳入结合模型,该模型表明其芳香部分与 claudin-5 细胞外结构域和相邻跨膜片段的高度保守残基相关联。我们的结果表明了以下作用模式:M01 优先与细胞外 claudin-5 结构域结合,从而削弱粘附细胞之间的反式相互作用。由于内化和转录下调,膜状 claudin-5 水平进一步降低,使小分子能够通过细胞旁路。总之,这里引入的第一个小分子是作为药物增强剂,它特异性地使 BBB 通透足够长的时间,以允许神经药物进入大脑。