XiaoMi-AI文件搜索系统
World File Search Systemenhancer

DNA转录的神秘和美丽-Iris Publishers
图9:哺乳动物转录的调节。通过形成染色体环的形成,使DNA的活跃增强子调节区域可与靶基因的启动子DNA区域相互作用。这可以通过RNA聚合酶II(RNAP II)启动Messenger RNA(mRNA)合成,并在基因的转录开始位点与启动子结合。通过锚定在增强子上的一个结构蛋白稳定环,一个固定在启动子上的蛋白质,将这些蛋白固定在启动子上,并连接到形成二聚体(红色锯齿形)。特定的调节转录因子与增强子上的DNA序列基序结合。一般转录因子与启动子结合。当转录因子被信号激活时(此处指示为增强子上的转录因子上的小红色星形所示的磷酸化),增强子被激活,现在可以激活其目标启动子。通过绑定的RNAP IIS在相反的方向上在DNA的每条链上转录活性增强子。介体(一个由相互作用结构中约26个蛋白质组成的复合物)将调节信号从增强子DNA结合的转录因子传达给启动子。
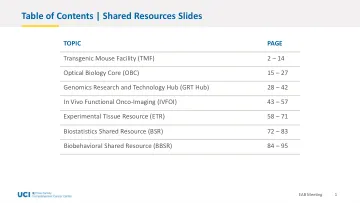
目录 | 共享资源幻灯片
• 由于缺乏有效的体内模型,与人类遗传病相关的非编码变异的功能表征仍然具有挑战性。 • Dual-enSERT-2 是一种强大的基于 CRISPR 的双色荧光报告系统,可快速定量分析活体 G0 第一代转基因小鼠中的增强子等位基因活性。 • Dual-enSERT 可与单细胞转录组学相结合,以细胞分辨率表征变异增强子等位基因活性,揭示与致病增强子失调有关的候选分子通路(例如介导神经胶质瘤形成的 IDH1 增强子变异)。 • TMF 对小鼠受精卵和胚胎进行显微注射和电穿孔,并将其转移到假孕受体小鼠中供 Kvon 实验室使用。

3D基因组和预测基因调控模型
• 转录调控由与启动子和增强子元件结合的转录因子 (TF) 协调 • 远端增强子可能距离启动子 >1Mb,通过染色质环路进行物理相互作用 • 1D 表观基因组数据(染色质可及性、组蛋白标记)映射候选增强子元件的存在但不映射它们的连接性

新英格兰生物实验室分析证书
在 250 ng vTPA(截短组织型纤溶酶原激活剂)模板质粒 DNA、1 µl 增强剂 1、1 µl 增强剂 2 和 20 单位 RNase 抑制剂(含有 PURExpress® 体外蛋白质合成试剂盒的成分)存在下进行 25 µl 反应,在 37°C 下孵育 2 小时,通过 SDS-PAGE 和考马斯亮蓝检测测定,可得到预期的 40 kDa 产物。

缺乏 Sry 的 Amami 棘鼠的哺乳动物性染色体的周转是由于雄性特异性的 Sox9 上调
哺乳动物的性染色体是高度保守的,性别由 Y 染色体上的 SRY 决定。两种特殊的啮齿动物群(其中一些物种缺少 Y 染色体和 Sry)为我们了解新的性基因如何产生并取代 Sry ,从而导致性染色体周转提供了见解。然而,30 多年的深入研究未能揭示这两个谱系中新的性基因的身份。我们在此报告在奄美刺鼠 Tokudaia osim- ensis 中发现了雄性特异性的 Sox9 增强子重复,这种大鼠的雄性和雌性都只有一条 X 染色体(XO/XO),而 Y 染色体和 Sry 完全丢失。我们进行了全面的调查以检测刺鼠中性别特异性的基因组区域。性别相关的基因组差异仅限于雄性特异性的 17 kb 单位重复,该重复位于常染色体上 Sox9 上游 430 kb 处。使用雄性刺鼠细胞进行的 Hi-C 分析表明,重复区域具有与 Sox9 的潜在染色质相互作用。重复单元含有一个与小鼠增强子 14 (Enh14) 同源的 1,262 bp 元件,Enh14 是一种候选 Sox9 增强子,在小鼠中功能冗余。转基因报告小鼠表明,刺鼠 Enh14 可作为小鼠胚胎睾丸增强子发挥作用。用重复的刺鼠 Enh14 替换 Enh14 的 XX 小鼠的胚胎生殖腺显示 Sox9 表达增加,Foxl2 表达减少。我们提出,这种 Sox9 增强子的雄性特异性重复取代了 Sry 功能,从而定义了刺鼠中的一种新型 Y 染色体。

mecom是通过控制急性髓样白血病
增强子易位,由于3q26重排,在急性髓样白血病(AML)的侵略性亚型中驱动了脱皮的MECOM表达。使用内源性生长素诱导的脱基龙直接耗尽MECOM,立即上调髓样分化因子CEBPA的表达。MECOM耗竭也伴随着干细胞的严重丧失和分化的增加。mecom通过与 +42KB CEBPA增强子结合而产生抑制作用,这是中性粒细胞发育必不可少的基因。这部分取决于MECOM及其共抑制器CTBP2之间的相互作用。我们证明CEBPA过表达可以绕过MECOM介导的分化块。此外,通过增强劫持的AML MECOM过表达的AML患者显着降低了CEBPA。我们的研究直接连接了两个主要参与者Meeloid Transformation Mecom和Cebpa,它提供了对MECOM通过灭活Cebpa将干细胞状态保持在AML独特亚型的机制的洞察力。

事件议程
Exploring cross-trait genetic architectures: the BIGA platform 1:30 PM - 3:15 PM Speaker: Bingxin Zhao Genetic imputation models from single-cell and bulk-tissue transcriptomics for investigating the genetic architecture of complex traits 1:30 PM - 3:15 PM Speaker: Eric Gamazon Mapping Enhancer Networks to Human Disease Genetics 1:30 PM - 3:15 PM Speaker: Xiang Zhu大规模特质的某些应用与基因分型个体和GWAS摘要数据的某些应用1:30 pm -3:15 PM扬声器:WEI PAN

发育生物学学会第83届年会...
摘要书籍程序摘要#1探索阴影增强剂建筑的功能意义Jillian Ness,Yu Wang,Christian Mei,Renata Serio,Renata Serio,Zeba Wunderlich Boston University,美国,美国许多发育基因都受到一套看似冗余的增强剂的调节,这些增强剂可以推动重叠的Spatiotemporpormoral模式。这些多增强器系统被称为阴影增强剂。阴影增强子可以缓冲遗传和环境应力以驱动正常的基因表达模式。使用果蝇胚作为模型,我们表明,前后图案基因的阴影增强子通过结合不同的输入转录因子的不同集来驱动一致的基因表达模式。尽管如此,尚不清楚为什么阴影增强子转录因子结合位点分布在多个增强器上,而不是在单个增强器中。可以想象,阴影增强子使用的机制可以编码为单个增强剂。我们已经生成了增强剂记者,其中消除了阴影增强剂之间的内源性DNA以做出“挤压”配置。令人惊讶的是,我们发现毛压和内源间隔增强子之间的RNA模式,水平和动力学几乎没有差异,这表明阴影功能不需要内源性间距。我们正在构建各个长度的合成污垢增强子,以测试squish增强子保留功能的较低尺寸限制。要理解引起阴影增强子的进化动力学,我们已经使用生物信息学来识别发育阴影增强子集的起源。星期四,我们发现重复事件和可转座元素似乎是果蝇中相对较小的(<10%)的发育阴影增强子出生来源,与我们在小鼠基因组中的发现相比(〜30%)。这些数据表明,合并的单个增强剂可以与不同的阴影增强子相比功能,而不会在压力下损害忠诚度。我们将探测创造和维持阴影增强子的进化动力学,以使其在动物发育中的普遍作用合理化。由NIH授予R01HD095246资助的工作。

停止使用 COVID-19 mRNA 疫苗
卫生局长概述了对已获批准的辉瑞和 Moderna COVID-19 mRNA 疫苗中核酸污染物的担忧,特别是在存在脂质纳米颗粒复合物和猿猴病毒 40 (SV40) 启动子/增强子 DNA 的情况下。脂质纳米颗粒是将 COVID-19 疫苗中的 mRNA 递送到人体细胞的有效载体,因此可能也是将污染物 DNA 递送到人体细胞的同样有效的载体。SV40 启动子/增强子 DNA 的存在也可能带来 DNA 整合到人体细胞中的独特且更高的风险。
