XiaoMi-AI文件搜索系统
World File Search Systemenhancer

KDM1A 维持转录增强子的全基因组稳态
转录增强子能够对后生动物的基因表达进行精确的时空控制。组蛋白 H3 赖氨酸 4 (H3K4me1) 的单甲基化富集是转录增强子的主要染色质特征。赖氨酸 (K) 特异性脱甲基酶 1A (KDM1A,也称为 LSD1) 是一种 H3K4me2/me1 脱甲基酶,可在小鼠胚胎干细胞 (mESC) 分化过程中使干细胞增强子失活。然而,其在未分化 mESC 中的作用仍不清楚。在这里,我们表明 KDM1A 在未分化和谱系定向细胞中都积极维持最佳增强子状态。KDM1A 占据了未分化 mESC 中的大部分增强子。增强子处的 KDM1A 水平与其底物 H3K4me2、H3K27ac 和增强子处的转录呈现明显的正相关性。在缺乏 Kdm1a 的 mESC 中,这些增强子中的大部分获得了额外的 H3K4 甲基化,同时伴有 H3K27 乙酰化增加以及增强子 RNA (eRNA) 和靶基因表达增加。在有丝分裂后的神经元中,KDM1A 的缺失会导致神经元活动依赖性增强子和基因的过早激活。总之,这些结果表明 KDM1A 是一种多功能的增强子调节器,并充当变阻器,通过平衡增强子处的 H3K4 甲基化来维持最佳增强子活性。

p13-推定的人类增强子在大脑进化中的作用...
摘要:转录因子SOX2调节了几种基因,这些基因在突变时会引起神经发育障碍(NDDS)。sox2与远端增强子结合,维持增强子促销相互作用网络的完整性,进而维持适当的基因表达水平。此外,我们在小鼠神经干细胞(未分化和分化为神经元/神经胶质中)中通过Chipseq和Chia-pet鉴定了约10000个“表观遗传增强子”及其与启动子的长距离相互作用。其中约有7500个在人类中是保守的,也称为人类交通远程相互作用区域(“ hmslris”)。其中许多与NDD相关的DNA序列变体重叠;通过我们的远程相互作用的地图,这些增强子相关的变体表明有助于NDD的新型候选基因。进一步,我们鉴定了微观恐惧症/植血性/嗜性症/coloboma(MAC)患者的微型缺失/重复,这些患者去除/重复的单个增强子(以野生型)与眼睛相关的基因相关,这表明这些增强剂的损失/增加可能会导致疾病。在一项平行研究中,我们确定了HMSLIS与某些人类加速区域(HAR)之间的36个重叠。Hars序列在物种之间是高度保守的,但在人类中显示出很大的变化。此外,hars在脑发育中涉及的基因附近富含,并且已显示出几种hars可以用人类特异性活动的变化编码转录增强子。我们专注于与HAR重叠的HMSLRI,并且与BTG1基因具有广泛的相互作用,这对于小鼠脑发育很重要。我们将研究连接到BTG1的增强子的体内功能,以及使用斑马鱼中的转基因增强剂测定法与与MAC相关的基因具有长距离相互作用的四个增强子。如果我们发现斑马鱼中的增强剂活性,我们可以分析HMSLRI BTG1中人类特异性取代(HSUB)的功能效应以及与MAC相关基因相关的HMSLRIS中的变体。
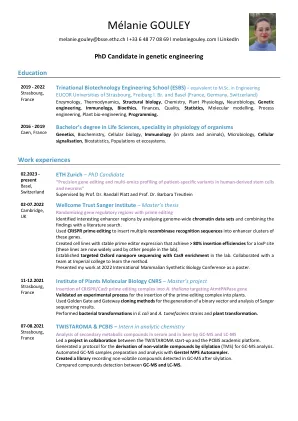
梅兰妮·古利
威康基金会桑格研究所 – 硕士论文 使用 prime editing 随机化基因调控区域 通过分析全基因组染色质数据集并将研究结果与文献检索相结合,确定了有趣的增强子区域。 使用 CRISPR prime editing 将多个重组酶识别序列插入这些基因的增强子簇中。 创建了具有稳定 prime editor 表达的细胞系,可实现 loxP 位点 80% 以上的插入效率(这些细胞系现在被实验室中的其他人广泛使用)。 在实验室中建立了具有 Cas9 富集的靶向牛津纳米孔测序。 与帝国理工学院的一个团队合作学习该方法。 在 2022 年国际哺乳动物合成生物学会议上以海报形式展示了我的工作。 2021 年 11 月 12 日,法国斯特拉斯堡

F631S Phusion™ Plus PCR 预混液
批次 数量 描述 2727858 2 x 1.25 mL Phusion™ Plus PCR Master Mix 2740002 1.25 mL Phusion™ GC 增强剂 2720703 2 x 1.25 mL 水,无核酸酶


增强子 - 基因在发育和疾病中的特异性
增强子控制整个发育过程中时空基因表达模式的建立。在过去的十年中,新技术的发展提高了我们根据其在同一拓扑结构域内共定位将增强子与目标基因联系起来的能力。然而,调节增强子如何专门激活某些基因而但在给定领域内的其他机制尚不清楚。在这篇综述中,我们讨论了对控制增强子特异性的因素的最新见解,包括增强子和启动子的遗传组成,增强子及其靶基因之间的线性和3D距离以及细胞型特异性染色质景观。我们还讨论如何阐明增强子特异性的分子原理可能有助于我们更好地理解和预测人类遗传,表观遗传和结构变异的病理后果。

附录8-新加坡
调节和功能遗传元件(例如启动子,增强子,限制酶位点,转基因和选择标记物)。信息包括但不限于病毒capsID的组成,包膜结构,分子量,粒径,糖基化位点,基因组的性质(单链,双链,DNA或RNA,DNA或RNA,每个颗粒的基因组的拷贝数),病毒载体的热门(例如, 病毒载体对特定宿主组织的特异性)。 •对于质粒载体,提供调节和功能遗传元件的示意图(例如) 启动子,增强子,限制酶位点,转基因和选择标记物)。 信息包括但不限于物理特性,生化特征,遗传标记和位置(例如) 插入的外源基因的质粒,偶发或染色体)。 •用于使用基因编辑技术,提供病毒载体对特定宿主组织的特异性)。•对于质粒载体,提供调节和功能遗传元件的示意图(例如启动子,增强子,限制酶位点,转基因和选择标记物)。信息包括但不限于物理特性,生化特征,遗传标记和位置(例如插入的外源基因的质粒,偶发或染色体)。 •用于使用基因编辑技术,提供插入的外源基因的质粒,偶发或染色体)。•用于使用基因编辑技术,提供
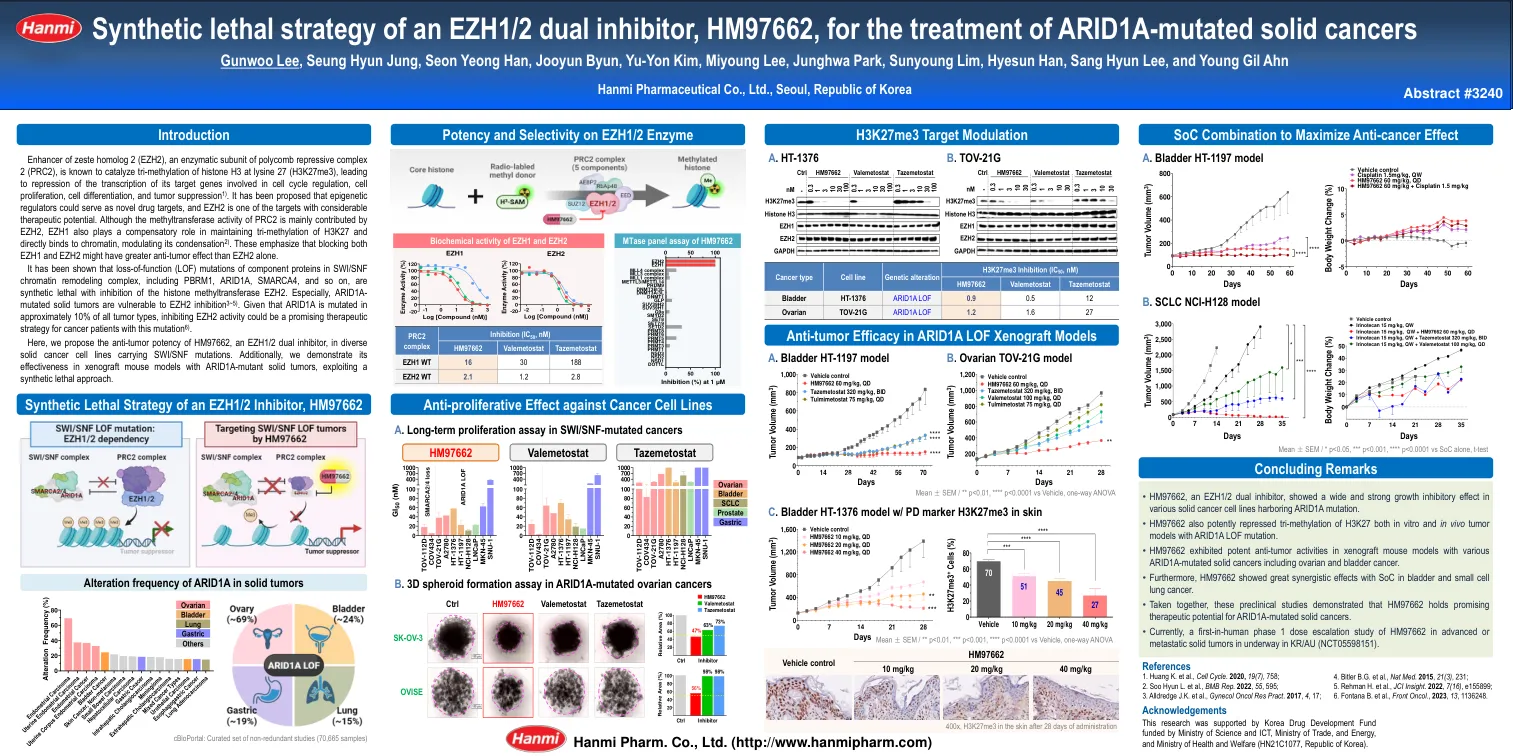
一种新颖且有效的EZH1/2双重抑制剂,HM97662表现出广泛的治疗潜力
Enhancer of zeste homolog 2 (EZH2), an enzymatic subunit of polycomb repressive complex 2 (PRC2), is known to catalyze tri-methylation of histone H3 at lysine 27 (H3K27me3), leading to repression of the transcription of its target genes involved in cell cycle regulation, cell proliferation, cell differentiation, and tumor suppression 1) .已经提出表观遗传调节剂可以用作新的药物靶标,而EZH2是具有巨大治疗潜力的靶标之一。尽管PRC2的甲基转移酶活性主要由EZH2贡献,但EZH1在维持H3K27的三甲基化方面也起着补偿性作用,并直接与染色质结合,调节其凝结2)。这些强调,与单独的EZH2相比,阻止EZH1和EZH2的抗肿瘤效应可能更大。

临床试验和产品的 ctgtp 的 cmc 要求 - ...
调控和功能性遗传元件(例如启动子、增强子、限制性酶切位点、转基因和选择标记)的示意图。信息包括但不限于病毒衣壳的组成、包膜结构、分子量、颗粒大小、糖基化位点、基因组的性质(单链、双链、DNA 或 RNA、每个颗粒的基因组拷贝数)、病毒载体的趋向性(例如病毒载体对特定宿主组织的特异性)。• 对于质粒载体,提供调控和功能性遗传元件(例如启动子、增强子、限制性酶切位点、转基因和选择标记)的示意图。信息包括但不限于插入的外来基因的物理特性、生化、生长特性、遗传标记和位置(例如在质粒上、游离型或染色体上)。• 对于基因编辑技术的使用,提供

PAX2构成了...
抽象协调的动物运动取决于功能前置体的发展。虽然早期的细胞效果确定过程是充分表征的,但对本体感受谱系中细胞的终末分化以及控制它们的遗传网络的终极分化知之甚少。在这项工作中,我们描述了一个基因调节网络,该网络由三个转化因子(Prospero(pros),D-PAX2和Delilah(DEI)组成,这决定了果蝇中的本体感受谱系中的两个替代分化程序。我们表明,D-Pax2和ProS分别通过激活和抑制DEI的转录来控制脊柱器官谱系中盖与scolopale细胞的分化。通常,D-PAX2激活了DEI在上限电池中的表达,但在Pros被共表达的Scolopale细胞中无法进行。我们进一步表明,D-Pax2和Pro通过262 bp核核定特异性增强剂对DEI转录产生影响,其中两个D-PAX2-和三个Pros结合位点实验鉴定出来。从蝇基因组中除去该增强子时,DEI的帽和韧带特异性表达丢失,从而导致核核器官功能的丧失和幼体幼虫的不良运动。因此,协调的幼虫运动取决于DEI增强子的活性,该活性同时整合了动作和抑制性输入,以生成功能性前置的器官。