XiaoMi-AI文件搜索系统
World File Search Systemgrnas

基于系统的基于图像的基于斑马鱼幼虫中复杂性状的遗传筛选
摘要具有数以千计的基因组关联研究对复杂特征鉴定的基因座,需要在体内模型系统中可靠,迅速推断大量候选基因的作用。基于F 0斑马鱼中的基于CRISPR/CAS9的功能屏幕代表这样的系统。然而,到目前为止使用的负面对照 - 包括加扰的指南RNA(GRNA),灭活的CAS9和假注射 - 不会引起与CRISPR/CAS9相同的细胞和有机反应,并且可能会加剧结论。在这里,我们表明,靶向KITA促进了成功的诱变,更高质量的成像数据以及病例和对照的有效分类的有效的光学预筛查。我们鉴定并测试了两个靶向具有类似高诱变效率和对色素作用的kita的GRNA,并且没有对心脏代谢性状的脱靶效应或主要影响。我们提出了几种方法,这些方法将得出有效的,公正的结论。

Invitrogen转染产品
CRISPR-CAS9是一种用于基因组编辑和工程的既定方法,包括在对特定基因组裂解事件不完善的修复后,基因表达敲除诱导。Invitrogen™Lentipool V2人CRISPR文库已开发用于功能丧失研究,以确定基因在调节细胞过程中的作用以及对化合物,药物或任何可能影响这些过程的任何扰动者的细胞反应。Lentipool V2人类CRISPR库中包含的CRISPR-CAS9指南RNA(GRNA)是基于专有GRNA设计算法设计的,该算法选择指南以最大程度地敲除效率而不牺牲特异性。每个库包含每个基因的四个序列验证的不同的GRNA矢量构建体,包装为慢病毒颗粒。Lentipool V2人CRISPR库由定制或预定义集合的grnas组成,以特定基因家族为目标,以慢病毒合并格式进行功能基因组学筛选,请参见第29页。

小麦 Sal1 基因家族的 CRISPR-Cas9 基因编辑
摘要:高度保守的 Sal1 编码一种双功能酶,具有肌醇多磷酸-1-磷酸酶和 3 ′ (2 ′),5 ′-二磷酸核苷酸酶活性,已被证明在被破坏时会改变植物的非生物胁迫耐受性。精确的基因编辑技术被用于在六倍体面包小麦中产生 Sal1 突变体。带有三个向导 RNA (gRNA) 的 CRISPR(成簇调节间隔短回文重复序列)Cas9 系统被用于灭活 Bobwhite 小麦基因组中的六个 Sal1 同源基因。所产生的所有 Sal1 基因均被禁用的突变小麦植株茎更细,生物量略有减少,在缺水条件下衰老更慢,但在干旱条件下产量没有提高。我们的结果表明,多重 gRNA 能够有效地对六倍体小麦中的 Sal1 基因家族进行靶向基因编辑。这些 Sal1 突变小麦植株将成为进一步研究该基因家族在小麦中功能的资源。

使用 CRISPR 条形码作为分子钟,以单细胞分辨率捕捉动态过程
图 1. a. 带有 poly-A 读数的动态条形码示意图。b. 实验装置的示意图。c. 基于突变特征的条形码比例,结合两个系统的数据:对具有完整 PAM 基序的原型间隔物进行编辑(活性);不存在 PAM 基序(非活性);和未切割的 gRNA(原始)。d. 不同 gRNA 中原始条形码随时间的比例。e. 考虑不同 gRNA 之间的错配、间隙和间隙延伸,条形码随时间的变化。f. 具有 21 bp 间隔物(左)或 26 bp 间隔物(右)的 gRNA 的原始条形码随时间的比例。箱线图按不同时间点的平均间隔物长度着色(Cas9 系统)。g. 原始核苷酸随时间变化的百分比,将间隔物相对于 PAM 序列对齐(Cas9 系统)。h。考虑到按 Cas9 版本分类的所有不同 gRNA,C>T 突变随时间变化的百分比。对于所有箱线图,箱线表示四分位距 (IQR),每个箱线内的水平线表示中位数。

MultiCrispr:用于数千个目标的主要编辑和平行定位的GRNA设计
针对编码基因组通过CRISPR/ CAS9技术引入核苷酸缺失/插入已成为一种标准程序。它迅速产生了多种方法,例如素数编辑,顶点接近标记或同源性修复,但是,支持生物信息学工具的支持落后于此。新的CRISPR/CAS9应用程序通常会重新征询特定的GRNA设计功能,并且通常缺少一种通用工具。在这里,我们介绍了R/生物导体工具MulticRispr,旨在设计单个grnas和复杂的grna libraries。包装易于使用;在效率和特定的效率上,检测,分数和锻炼;每个目标或CRISPR/CAS9序列可视化和聚集结果;最后返回GRNA的范围和序列。是通用的,多晶状体定义的,并实现了基因组算术框架,作为便利适应最近引入的技术的基础,例如素数编辑或尚未出现。其性能和设计构想(例如目标集) - 特定过滤渲染多晶层在处理类似筛选的方法时选择的工具。

crispr-tsko的金门装配
在第一步中,将六个金门入口向量合并为目标向量。有各种可以使用的金门目标向量,其中包含可以使用的不同植物和/或视觉标记物(请参阅补充数据集1中的金门目标矢量(CCDB +)1)。第一个入口向量(AB)包含组织特异性表达的启动子。第三个入口矢量(CD)包含核酸酶,可以与N末端(BC)或C末端标签(DE)结合使用。另外,如果不需要标签,则使用链接序列。第五入口矢量(EF)包含工厂终结器。选择的第六个黄金入口向量(FG)取决于最终目标。要克隆与一个或两个GRNA兼容的矢量,请使用未武装的GRNA进入矢量PGG-F-F-ATU6-26-AARI-AARI-AARI-G(请参阅补充数据集1中的未武装GRNA进入向量1)。要克隆与多个GRNA兼容的矢量,请使用可变的链接器PGG-F-a-aari-sacb-aari-g-g(请参阅补充数据集1中的可变链接器)。由于我们的克隆策略使用限制酶Bsai和Aari,因此要求所有向量都需要无BSAI和AARI-FIME(除了克隆位点)。
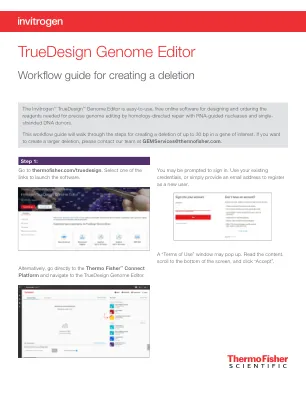
Truedesign基因组编辑器
要查看TALEN目标,请单击表的“ Talen”选项卡,并将显示每个Talen对的类似信息。TAL效应子核酸酶(TALEN)对。设计结果表中的绿色选中标记将指示推荐的技术。在Thermofisher.com/tal上了解有关Talen Technology的更多信息。
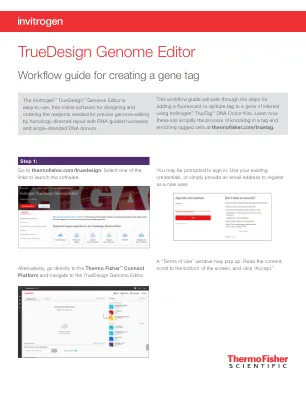
Truedesign基因组编辑器
要查看TALEN目标,请单击表的“ Talen”选项卡,并将显示每个Talen对的类似信息。TAL效应子核酸酶(TALEN)对。设计结果表中的绿色选中标记将指示推荐的技术。在Thermofisher.com/tal上了解有关Talen Technology的更多信息。
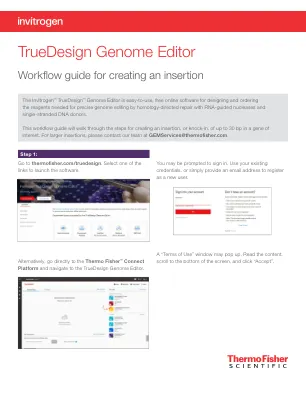
TrueDesign 基因组编辑器
要查看 TALEN 靶标,请单击表格中的 TALEN 选项卡,每个 TALEN 对都会显示类似的信息。当敲入位点 10 bp 范围内没有 PAM 位点,或者 gRNA 的效率和特异性不理想时,建议使用 TAL 效应核酸酶 (TALEN) 对。设计结果表中的绿色复选标记将指示推荐的技术。有关 TALEN 技术的更多信息,请访问 thermofisher.com/tal 。
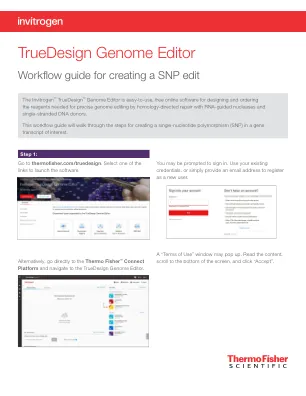
Truedesign基因组编辑器
要查看TALEN目标,请单击表的“ Talen”选项卡,并将显示每个Talen对的类似信息。TAL效应子核酸酶(TALEN)对。设计结果表中的绿色选中标记将指示推荐的技术。在Thermofisher.com/tal上了解有关Talen Technology的更多信息。