XiaoMi-AI文件搜索系统
World File Search Systemhomologous

猕猴顶上小叶 5 区 (PE) 细分的相应解剖结构揭示了与人类相似的连接模式
基于影像特征将动物脑作为跨物种研究的工具,可为揭示人类大脑的综合分析提供更多潜力。先前的研究表明,人类布罗德曼5区(BA5)和恒河猴的PE为同源区域,均参与手臂运动中触觉过程中的深度和方向信息处理。但最近的研究表明,BA5与PE并不同源,根据细胞构架,BA5被细分为三个不同的亚区域,PE可细分为PEl、PEla和PEm,BA5与PE之间各亚区域之间的物种同源关系尚不明确。同时,基于白质纤维束解剖连接对PE的细分需要更多的验证。本研究依据白质纤维束解剖连接对恒河猴的PE进行了细分。基于概率纤维追踪技术定义前侧和背侧两个PE亚区,最后针对BA5和PE亚区绘制具有预定义同源靶区的连通性指纹,揭示结构和功能特征,并给出识别出的同源对应关系。

气动参数智能算法...
本文提出了一种用于先进布局飞机大迎角风洞试验气动参数标定的智能算法,该算法基于同源比对与调优算法,可以有效提高风洞试验模型的精度。首先,在分析某先进布局缩比飞机大振荡风洞试验数据的基础上,建立了由静导数、动导数、旋转平衡导数组成的大迎角风洞试验模型。其次,为有效提高风洞试验模型的精度,提出了分层标定与智能算法相结合的大迎角同源比对修正思路。所提方法解决了先进布局飞机大迎角气动模型同源比对中结构复杂、数据量大、精度差的问题。最后基于MATLAB GUI设计了相应的比对界面软件,将提出的方法与思路融入其中,实现了先进布局飞机大迎角模拟飞行风洞试验气动参数的有效调整,为后续先进布局飞机大迎角飞行试验验证提供了可靠的工程技术手段。

粘蛋白指导通过环修复过程中的同源性搜索通过环和姐妹染色单体链接
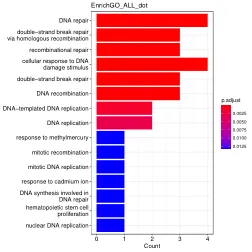
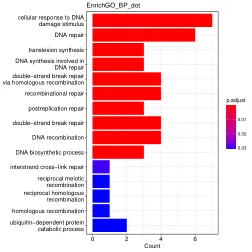
准确修复DNA双链断裂(DSB)对于基因组稳定性至关重要,并且有缺陷的修复是癌症等疾病的基础。同源重组使用完整的同源序列来忠实地恢复受损受损的DNA,但是损坏的DNA终止如何在包含数十亿个非同源碱基的基因组中找到同源位点,尚不清楚。在这里,我们介绍了姐妹孔C,这是一种高分辨率方法,用于绘制复制染色体中的分子内和转运相互作用。我们通过募集两个功能上不同的粘蛋白池来证明DSBS重塑染色体体系结构。环形成粘着蛋白积聚在巨型尺度范围内,以控制围绕破裂位点的拓扑关联结构域(TAD)内的同源性采样,而粘性粘着蛋白将浓缩的位点浓缩到蛋白质染色剂的链球末端。这种双重机制限制了同源性搜索空间,突出了染色体构象如何有助于保持基因组完整性。

使用crispr/ttlbcas12a用于planta基因靶向A. thaliana
CRISPR/CAS系统通过诱导特定位点DNA双链断裂(DSB)启用基因编辑。但是,诱导的修饰的性质高度取决于用于DNA DSB修复的机制。非同源末端连接(NHEJ)介导的靶向诱变是由CRISPR/CAS诱导的一种已经标准应用的工具,可以在特定的基因组位点导致各种不同种类的突变。尽管如此,使用同源供体序列的精确基因组修饰仍然具有挑战性。的应用取决于较不频繁的同源重组(HR)需要进一步改进,以创建一种有吸引力的植物应用工具。着眼于这个问题,我们开发了植物基因靶向(IPGT)系统,该系统基于同时切除稳定集成的同源供体序列和目标位点中DSB的诱导。近年来,增强了基因靶向(GT)频率的几种改进。在为IPGT链球菌CAS9(SP Cas9)和金黄色葡萄球菌Cas9(SA Cas9)成功地促进了,我们能够使用lachnospileceae cas12a(lb cas12a)进一步改善该系统,这也可以在T-rich区域进行切割。 最近,我们测试了IPGT的LB CAS12A(TT LB CAS12A)的改进,耐温度的版本,并能够进一步提高GT效率。 在这里,我们详细介绍了使用TT LB Cas12a详细介绍最近发布的IPGT系统的实验程序。 ©2020作者。,我们能够使用lachnospileceae cas12a(lb cas12a)进一步改善该系统,这也可以在T-rich区域进行切割。最近,我们测试了IPGT的LB CAS12A(TT LB CAS12A)的改进,耐温度的版本,并能够进一步提高GT效率。在这里,我们详细介绍了使用TT LB Cas12a详细介绍最近发布的IPGT系统的实验程序。©2020作者。

fignl1的分子基础从DNA和染色质分离rad51中
1。P. Baumann,F。E. Benson,S。C. West,Human Rad51蛋白在体外促进ATP依赖性同源配对和链转移反应。Cell 87,757-766(1996)。 2。 F. E. Benson,A。Stasiak,S。C。West,人类Rad51蛋白的纯化和表征,大肠杆菌的类似物。 EMBO J 13,5764-5771(1994)。 3。 y。 Sun,T。J. McCorvie,L。A. Yates,X。Zhang,同源重组的结构基础。 单元格。 mol。 生命科学。 77,3-18(2020)。 4。 D. K. Bishop,RecA同源物DMC1和RAD51相互作用,在减数分裂染色体突触之前形成多个核复合物。 Cell 79,1081-1092(1994)。 5。 A. Carver,X。Zhang,Rad51细丝动力学及其拮抗调节剂。 Semin Cell Dev Biol 113,3-13(2020)。 6。 Y. W. Chan,S。C. West,一种由同源重组产生的新的超级后期桥。 细胞周期17,2101-2109(2018)。 7。 A. Piazza,W。D. Wright,W.-D。海耶尔(Heyer),多侵略是诱导染色体重排的重组副产物。 单元格170,760-773.E715(2017)。 8。 K. Schlacher,H。Wu,M。Jasin,一种独特的复制叉保护途径将fanconi贫血肿瘤抑制剂连接到RAD51-BRCA1/2。 癌细胞22,106-116(2012)。 9。 S. Tye,G。E。Ronson,J。R。Morris,道路上的叉子:同源重组和停滞的复制叉保护部分。 Semin Cell Dev Biol 113,14-26(2021)。Cell 87,757-766(1996)。2。F. E. Benson,A。Stasiak,S。C。West,人类Rad51蛋白的纯化和表征,大肠杆菌的类似物。EMBO J 13,5764-5771(1994)。3。y。Sun,T。J. McCorvie,L。A. Yates,X。Zhang,同源重组的结构基础。单元格。mol。生命科学。77,3-18(2020)。 4。 D. K. Bishop,RecA同源物DMC1和RAD51相互作用,在减数分裂染色体突触之前形成多个核复合物。 Cell 79,1081-1092(1994)。 5。 A. Carver,X。Zhang,Rad51细丝动力学及其拮抗调节剂。 Semin Cell Dev Biol 113,3-13(2020)。 6。 Y. W. Chan,S。C. West,一种由同源重组产生的新的超级后期桥。 细胞周期17,2101-2109(2018)。 7。 A. Piazza,W。D. Wright,W.-D。海耶尔(Heyer),多侵略是诱导染色体重排的重组副产物。 单元格170,760-773.E715(2017)。 8。 K. Schlacher,H。Wu,M。Jasin,一种独特的复制叉保护途径将fanconi贫血肿瘤抑制剂连接到RAD51-BRCA1/2。 癌细胞22,106-116(2012)。 9。 S. Tye,G。E。Ronson,J。R。Morris,道路上的叉子:同源重组和停滞的复制叉保护部分。 Semin Cell Dev Biol 113,14-26(2021)。77,3-18(2020)。4。D. K. Bishop,RecA同源物DMC1和RAD51相互作用,在减数分裂染色体突触之前形成多个核复合物。Cell 79,1081-1092(1994)。 5。 A. Carver,X。Zhang,Rad51细丝动力学及其拮抗调节剂。 Semin Cell Dev Biol 113,3-13(2020)。 6。 Y. W. Chan,S。C. West,一种由同源重组产生的新的超级后期桥。 细胞周期17,2101-2109(2018)。 7。 A. Piazza,W。D. Wright,W.-D。海耶尔(Heyer),多侵略是诱导染色体重排的重组副产物。 单元格170,760-773.E715(2017)。 8。 K. Schlacher,H。Wu,M。Jasin,一种独特的复制叉保护途径将fanconi贫血肿瘤抑制剂连接到RAD51-BRCA1/2。 癌细胞22,106-116(2012)。 9。 S. Tye,G。E。Ronson,J。R。Morris,道路上的叉子:同源重组和停滞的复制叉保护部分。 Semin Cell Dev Biol 113,14-26(2021)。Cell 79,1081-1092(1994)。5。A.Carver,X。Zhang,Rad51细丝动力学及其拮抗调节剂。Semin Cell Dev Biol 113,3-13(2020)。6。Y. W. Chan,S。C. West,一种由同源重组产生的新的超级后期桥。细胞周期17,2101-2109(2018)。7。A. Piazza,W。D. Wright,W.-D。海耶尔(Heyer),多侵略是诱导染色体重排的重组副产物。 单元格170,760-773.E715(2017)。 8。 K. Schlacher,H。Wu,M。Jasin,一种独特的复制叉保护途径将fanconi贫血肿瘤抑制剂连接到RAD51-BRCA1/2。 癌细胞22,106-116(2012)。 9。 S. Tye,G。E。Ronson,J。R。Morris,道路上的叉子:同源重组和停滞的复制叉保护部分。 Semin Cell Dev Biol 113,14-26(2021)。A. Piazza,W。D. Wright,W.-D。海耶尔(Heyer),多侵略是诱导染色体重排的重组副产物。单元格170,760-773.E715(2017)。8。K. Schlacher,H。Wu,M。Jasin,一种独特的复制叉保护途径将fanconi贫血肿瘤抑制剂连接到RAD51-BRCA1/2。癌细胞22,106-116(2012)。9。S. Tye,G。E。Ronson,J。R。Morris,道路上的叉子:同源重组和停滞的复制叉保护部分。Semin Cell Dev Biol 113,14-26(2021)。10。H. L. Klein,Rad51过表达对正常和肿瘤细胞的后果。DNA修复(AMST)7,686-693(2008)。11。R。B. Jensen,A。Carreira,S。C. Kowalczykowski,纯化的人BRCA2刺激了Rad51介导的重组。自然467,678-683(2010)。12。L. A. Greenhough等。,RAD51B – RAD51C – RAD51D -XRCC2肿瘤抑制剂的结构和功能。自然619,650-657(2023)。13。Y. Rawal等。,在同源重组中对BCDX2复杂功能的结构见解。自然619,640-649(2023)。14。E. Antony等。 ,SRS2通过蛋白质 - 蛋白质相互作用触发ATP周转和RAD51与DNA解离的蛋白质蛋白质相互作用来解散RAD51丝。 mol Cell 35,105-115(2009)。 15。 J. Simandlova等。 ,FBH1解旋酶在体外破坏RAD51丝,并调节哺乳动物细胞中的同源重组*。 生物学杂志288,34168-34180(2013)。 16。 J. D. Ward等。 ,重叠的机制促进了减数分裂双链破裂修复期间突触后RAD-51细丝拆卸。 mol细胞37,259-272(2010)。 17。 M. Ito等。 ,Fignl1 AAA+ ATPase重塑了舒适性DNA复制和减数分裂重组中的RAD51和DMC1丝。 nat。 社区。 14,6857(2023)。 18。 J. Yuan,J。Chen,有效的同源重组修复需要含Fignl1的蛋白质复合物。 proc。E. Antony等。,SRS2通过蛋白质 - 蛋白质相互作用触发ATP周转和RAD51与DNA解离的蛋白质蛋白质相互作用来解散RAD51丝。mol Cell 35,105-115(2009)。15。J. Simandlova等。,FBH1解旋酶在体外破坏RAD51丝,并调节哺乳动物细胞中的同源重组*。生物学杂志288,34168-34180(2013)。16。J. D. Ward等。 ,重叠的机制促进了减数分裂双链破裂修复期间突触后RAD-51细丝拆卸。 mol细胞37,259-272(2010)。 17。 M. Ito等。 ,Fignl1 AAA+ ATPase重塑了舒适性DNA复制和减数分裂重组中的RAD51和DMC1丝。 nat。 社区。 14,6857(2023)。 18。 J. Yuan,J。Chen,有效的同源重组修复需要含Fignl1的蛋白质复合物。 proc。J. D. Ward等。,重叠的机制促进了减数分裂双链破裂修复期间突触后RAD-51细丝拆卸。mol细胞37,259-272(2010)。17。M. Ito等。,Fignl1 AAA+ ATPase重塑了舒适性DNA复制和减数分裂重组中的RAD51和DMC1丝。nat。社区。14,6857(2023)。18。J. Yuan,J。Chen,有效的同源重组修复需要含Fignl1的蛋白质复合物。proc。natl。学院。SCI。 110,10640-10645(2013)。 19。 Q. Zhang等。 ,flip-fignl1复合物调节在同源重组和复制叉重新启动中RAD51/DMC1的解离。 核酸Res 43,GKAD596(2023)。SCI。110,10640-10645(2013)。19。Q. Zhang等。 ,flip-fignl1复合物调节在同源重组和复制叉重新启动中RAD51/DMC1的解离。 核酸Res 43,GKAD596(2023)。Q. Zhang等。,flip-fignl1复合物调节在同源重组和复制叉重新启动中RAD51/DMC1的解离。核酸Res 43,GKAD596(2023)。

恶性疟原虫的血清型由免疫血清抑制体外生长*
体外生长抑制测定法用于检测恶性疟原虫菌株之间的抗原差异。猫头鹰猴的免疫。营养不良的猴子血清用于抑制八种恶性疟原虫菌株的体外生长。抑制是同源营地菌株的最大抑制作用(平均抑制100 mL/升cAMP-免疫血清)。其他四种菌株被较小程度抑制,三种菌株(FCR-3/FMG,FVO和Smith)在浓度高达400英里/升时并未受到cAMP免疫血清的显着抑制。fcr-3/fmg-rimmune血清,浓度为50 ml!升引起对FCR-3/FMG菌株的显着抑制,而不是cAMP菌株。因此,CAMP和FCR-3/FMG菌株似乎具有不同的抗原决定因素,而这些决定因素是同源性的,但不具有异源,抗血清的。通过免疫血清抑制体外生长可能对肺炎疟原虫的血清分型很有用,并且可能在选择菌株中应用于纳入疟疾疫苗。

原创文章 ICLE 遗传因素对人类大脑连接影响的半球间关系
要了解人类大脑线路中半球间差异和共性/耦合的起源,确定左右半球同源区域间连接是如何由遗传决定和关联的至关重要。为此,在本研究中,我们用高质量的扩散磁共振成像纤维束成像分析了人类双胞胎和家系样本,并估计了同源左右白质 (WM) 连接的遗传性和遗传相关性。结果表明,两个半球之间 WM 连接的遗传性相似且耦合,并且同源 WM 连接的遗传因素(即半球间遗传相关性)的重叠程度在整个大脑中差异很大:从完全重叠到完全不重叠。特别地,皮层下 WM 连接的遗传性明显强于皮层 WM 连接,并且遗传因素在半球间完全重叠的机会更高。此外,长距离连接的遗传性和半球间遗传相关性比短距离连接更强。这些发现突出了 WM 连接及其半球间关系背后的遗传学决定因素,并深入了解了健康和疾病状态下 WM 连接不对称的遗传基础。

通过体细胞核转移(SCNT)
这种SCNT卵母细胞的人工激活导致细胞分裂和染色体分离为伪极性体,并以70%的效率下的二核原体。与正常二倍体(n = 46)数量相比,极性体和Zygotes中单个染色体的下一代测序表明,染色体的数量降低了近一半(n = 19)(n = 19)。同源对的全面测序表明,平均将23对同源对的一半(n = 11)正确分离为极体和合子,而剩余的染色体对保持在一起,导致了肾上变。未检测到体细胞同源物之间的重组证据。