XiaoMi-AI文件搜索系统
World File Search Systempolyadenylation

RNA 编辑内含子 ENCO 转录本...
在前面的单元中,您已经学习了真核生物中 mRNA 的加工过程。主要的加工反应是 3' 端的聚腺苷酸化,这是加工事件,还有另一种加工,以转录后过程的形式被发现,转录前但在翻译之前,成熟 RNA 的核苷酸序列在外显子中编码的核苷酸序列不同 D 因此,RNA 编辑的定义可以是 d 哪些细胞可以在其转录本之后进行离散的 RNA 分子的生物合成替代蛋白质 p RNA 编辑过程酶

应用程序:4805 Deep Genomics
自 2022 年以来,Deep Genomics 已成为基因组研究和开发的先驱。BigRNA 以任何基因序列作为输入,然后可以准确预测数千种不同的分子生物学结果,从而能够在各种疾病、组织类型和遗传结构中发现靶点、疾病机制和 RNA 疗法。BigRNA 还能够从物种、组织、细胞模型、变体、基因、寡核苷酸和编辑等方面快速探索 RNA 生物学,包括剪接、多聚腺苷酸化、蛋白质/microRNA 结合、体外验证和对多种治疗方式的支持的机制。

TrkB.T1 作为慢性疼痛的治疗靶点
原肌球蛋白相关受体激酶 B (TrkB) 是脑源性神经营养因子 (BDNF) 的受体;其信号传导通过激活几个下游级联,有助于神经元存活、可塑性、分化和生长。缺乏细胞内激酶结构域的截短异构体 (TrkB.T1) 的过度表达与慢性疼痛的发展和持续有关。已发表的数据显示,小鼠模型中的 TrkB.T1 敲除可恢复运动功能并减轻脊髓损伤后的疼痛。我们项目的目标是确定抑制 TrkB.T1 表达的小分子作为慢性疼痛的潜在疗法,重点关注两种调节机制:(1) TrkB 前 mRNA 的差异转录后加工,以及 (2) 通过其 mRNA 的 3' 非翻译区 (3'UTR) 对 TrkB.T1 表达的转录后调节。对于第一点,我们假设两种主要 TrkB 亚型的比例主要受上游 (T1) pA 位点的切割和多聚腺苷酸化 (pA) 位点识别控制,因此抑制该位点 3'-加工的药物应能抑制 TrkB.T1 合成。对于第二点,我们假设 TrkB.T1 mRNA 3'UTR 包含调节序列,这些序列的功能可通过操纵关键反式因子的功能、表达或 RNA 结合活性的化合物进行调节,从而通过加速 TrkB.T1 mRNA 的衰变和/或抑制其翻译来抑制 TrkB.T1 的产生。对于每种机制,我们开发了独立的活细胞高通量筛选 (HTS) 检测方法,以识别可以 (1) 阻断 TrkB.T1 pA 位点的 3'-切割和多聚腺苷酸化,或 (2) 通过 TrkB.T1 mRNA 3'UTR 抑制基因表达的小分子。利用这些发现,我们旨在发现一种或多种能够抑制 TrKB.T1 表达的新药物,这些药物可在慢性疼痛的小鼠模型中作为新型镇痛药进行测试。
Mukherjee_ASGCT USH2A 海报_最终版
图 1:EDIT-102 的作用机制。*USH2A 基因中的 Ex13 代表导致 IRD 的任何外显子 13 突变,包括 c.2299delG。EDIT-102 编码人类 U6 启动子、向导 RNA(gRNA;RSQ9145 和 RSQ9265)、hGRK1 启动子、SV40(猿猴病毒 40)SD/SA(剪接供体/剪接受体)序列元素、NLS(核定位序列)、Sa(金黄色葡萄球菌)Cas9(CRISPR 相关蛋白 9)和 pA(多聚腺苷酸化信号)。EDIT-102 在 USH2A 外显子 13 的两侧进行编辑,导致外显子 13 从基因组和 mRNA 中去除,从而产生缺乏氨基酸 723-936 的功能性 Usherin 蛋白。

重新审查合成ORF序列对工程线路1逆转录的影响
A.从左到右:每个构造的示意图,一个来自逆转录术539分析的代表性井以及相对于SML1的逆转录效率。L1SPA_DBL_SMBB包含L1SPA_DOBLE的ORF1和540 ORF2序列,在ORF1P启动密码子(如SML1),541),541和SML1结构的删除3'UTR上具有Kozak共识。Restore_3'utr等于l1spa_dbl_smbb,但完整的L1SPA 3'UTR恢复了Neo Cassette的上游542。grr_after_neo等效于543 L1SPA_DBL_SMBB,但与Neo 544盒的下游放置的L1SPA 3'UTR的G-RICH区域,并删除了人类L1 Polyadyenylation信号。直方图显示了三个545生物复制测定的平均值,每个测定法包括每个构造的三个技术重复。三角形,正方形,546和钻石形状表示每个生物学重复的个体值,误差线547代表生物学重复之间的标准偏差。548
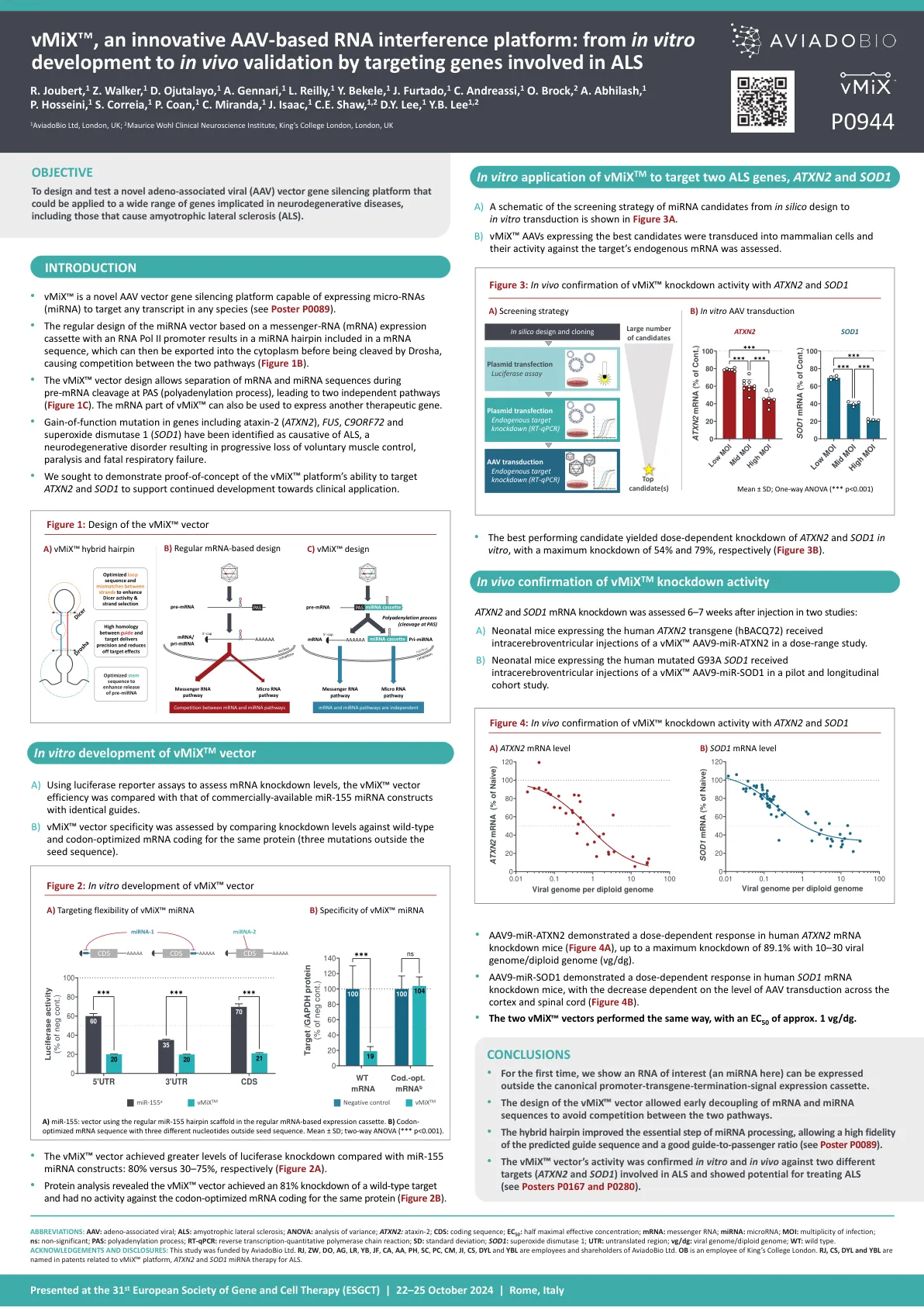
vmix,一种创新的基于AAV的RNA干扰平台
缩写:AAV:腺相关病毒; ALS:肌萎缩性侧索硬化;方差分析:方差分析; ATXN2:ataxin-2; CD:编码序列; EC 50:最大有效浓度的一半; mRNA:Messenger RNA; mirna:microRNA; MOI:感染的多样性; NS:不重要; PAS:聚腺苷酸化过程; RT-QPCR:逆转录 - 定量聚合酶链反应; SD:标准偏差; SOD1:超氧化物歧化酶1; UTR:未翻译的区域; VG/DG:病毒基因组/二倍体基因组; WT:野生型。致谢和披露:这项研究由Aviadobio Ltd. RJ,ZW,DO,AG,LR,YB,JF,CA,CA,AA,PH,SC,PC,PC,CM,JI,JI,CS,CS,CS,DYL和YBL是Aviadobio Ltd. OB的雇员和股东。rj,CS,dyl和ybl在与VMIX相关的专利中命名

cv_silvia barabino btbs unimib -milano -bicocca
近年来,我的研究小组的工作集中在与生理和病理条件下的MRNA剪接和聚腺苷酸化有关的一系列项目上。其中一个项目发现了mRNA输出和组蛋白PREMRNA的哺乳动物3'末端处理因子CF I的新功能。最近,我们表明线粒体应力会影响基因组大规模的替代剪接,因此可能有助于神经退行性疾病的发病机理。目前,我们专注于三个主要研究线。第一个集中在SRPK2上,SRPK2是一种针对SR(丝氨酸/精氨酸富域)剪接因子家族的蛋白激酶。我们目前正在研究DNA损伤通过SRPK2活性的调节影响替代剪接的机制。第二个项目涉及将选择替代外显子选择与转录的机制。具体而言,我们正在研究梵天是哺乳动物SWI/SNF染色质复合物的组成部分,如何通过与聚烯基化机制的组件相互作用来促进替代末端外显子的选择。最近,我们在肌萎缩性侧索硬化症的细胞和小鼠模型中启动了miRNA表达的表征。并行,我们目前正在研究RNA结合蛋白在基因组稳定性和神经变性中的作用。努力和激发年轻研究人员的能力。能够建立新的跨学科方法。简历生于索伦戈(CH),1963年7月9日。为了解决所有这些问题,我们使用了生化和分子方法的组合,例如CRIPR/CAS9基因组编辑,显微镜,体外细胞培养系统(包括IPSCS划分的细胞TYE)和转基因小鼠模型。已在实验室接受了20多名本科生和10名前后研究员的培训,其中许多人从事学术研究中的成功职业。我们一直与意大利和国外的许多研究小组合作。这包括由瑞士国家科学基金会资助的大型跨学科网络项目。1983-1987硕士学位。帕维亚大学

催化无活性 CRISPR 的新兴应用-...
摘要 rRNA、tRNA、mRNA和非编码RNA等不同类型RNA的转录后调控广泛参与生理和疾病的发生发展。mRNA作为基因与蛋白质之间的中间产物,其可变剪接、可变多聚腺苷酸化和修饰等转录后调控影响其编码蛋白的表达和功能。然而,由于缺乏合适的RNA工程平台,RNA转录后调控的功能意义和治疗潜力尚未得到充分研究。2015年一种特异性靶向RNA模板的新型CRISPR-Cas系统CRISPR-Cas13的发现,赋予了CRISPR以高特异性靶向和编辑RNA的新作用,在一定程度上开启了RNA操作的新时代。本综述将总结催化无活性的CRISPR-Cas13系统(CRISPR-dCas13)在mRNA工程中的新兴应用,并强调CRISPR-dCas13系统在其他RNA修饰调控中的前景及其治疗潜力。

使用合成 RNA 作为 RNA 直接测序中 poly(A) 长度推断的基准
多聚腺苷酸化是一个动态过程,在细胞生理学中非常重要。Oxford Nanopore Technologies 的直接 RNA 测序提供了一种对全长 RNA 分子进行测序以及对转录组和表观转录组进行分析的策略。目前有几种可用于 poly(A) 尾长估计的工具,包括 tailfindr 和 nanopolish 等成熟工具,以及两种较新的深度学习模型:Dorado 和 BoostNano。然而,这些工具的准确性与金标准数据集的基准测试有限。在本文中,我们使用合成 RNA 标准(Sequins)评估了四种 poly(A) 估计工具,这些标准具有已知的 poly(A) 尾长,并提供了一种衡量 poly(A) 尾长估计准确性的有效方法。所有四种工具生成的平均尾长估计值都在正确值的 12% 以内。总体而言,由于 Dorado 运行时间相对较快、变异系数低并且易于使用且可与碱基调用集成,因此被推荐作为首选方法。

揭示神经元分化过程中 RNA 异构体变化的动态和后果
静态基因表达程序已在干细胞和成熟人类细胞中得到广泛表征。然而,在细胞分化过程中,RNA 异构体随细胞状态转变而变化的动态、决定因素和功能后果在很大程度上仍不清楚。在这里,我们建立了一个改进的体外人类神经发生模型,该模型适用于全系统的基因表达分析。我们的多组学分析表明,细胞形态的显著改变与 RNA 异构体表达的广泛变化密切相关。我们的方法确定了在不同分化阶段表达的数千种新的 RNA 异构体。RNA 异构体主要来自外显子跳跃和人类神经发生过程中转录起始和多聚腺苷酸化位点的替代使用。转录异构体的变化可以重塑蛋白质异构体的身份和功能。最后,我们的研究确定了一组 RNA 结合蛋白是分化阶段特异性整体异构体变化的潜在决定因素。这项工作支持了神经发生过程中状态转变背后的受调控异构体变化的观点。