XiaoMi-AI文件搜索系统
World File Search Systemsecretion

果蝇中异二聚化依赖的 BMP 分泌
摘要 组合信号是指导情境相关细胞行为的关键。在胚胎发育、成体稳态和疾病期间,骨形态发生蛋白 (BMP) 充当二聚体来指导特定的细胞反应。BMP 配体可以形成同二聚体或异二聚体;然而,获得每种形式的内源性定位和功能的直接证据已被证明具有挑战性。在这里,我们利用精确的基因组编辑和通过蛋白质结合剂进行的直接蛋白质操作来剖析果蝇翅成虫盘中 BMP 同二聚体和异二聚体的存在和功能相关性。这种方法原位揭示了 Dpp (BMP2/4)/Gbb (BMP5/6/7/8) 异二聚体的存在。我们发现,尽管 Gbb 由翅成虫盘的所有细胞产生,但仅由表达 Dpp 的细胞分泌。 Dpp 和 Gbb 形成异二聚体的梯度,而在内源性生理条件下,Dpp 和 Gbb 同二聚体均不明显。我们发现异二聚体的形成对于在发育中的翅膀中获得最佳信号传导和长距离 BMP 分布至关重要。这些结果表明 Dpp/Gbb 异二聚体是上皮模式形成和生长所需的活性信号。

肠促胰岛素分泌的细胞机制
位于胃肠道上皮的肠内分泌细胞感知不同的营养物质/腔内内容物,从而触发各种肠道激素的分泌,这些激素在葡萄糖稳态和食欲调节中发挥不同的作用。肠促胰岛素激素胰高血糖素样肽-1 (GLP-1) 和葡萄糖依赖性促胰岛素多肽 (GIP) 在营养诱导下从肠道分泌后参与调节胰岛素分泌、食欲、食物摄入和体重。GLP-1 类似物已被开发并用于治疗 2 型糖尿病和肥胖症。调节内源性肠道激素的释放可能是治疗肥胖症和 2 型糖尿病的一种有前途的方法,无需手术。因此,本综述将重点介绍当前对肠道激素分泌细胞机制的理解。控制激素分泌的机制取决于刺激的性质,涉及多种信号通路,包括离子通道、营养转运蛋白和 G 蛋白偶联受体。

单氟化促进了α-突触核蛋白的分泌错误折叠相关的分泌。 美国多发性骨髓瘤患者产生的医疗费用(TCE)在美国
压力细胞秘密错误折叠的蛋白质,但是尚不清楚UCP中错误折叠的蛋白质的靶向错误。在这里,我们报告说,错误折叠的UCPS客户端会通过称为泛素蛋白 - 折叠式修饰剂1(UFM1)的泛素样蛋白进行修饰。Using α -synuclein ( α -Syn) as a UcPS model, we show that mutating the UFMylation sites in α -Syn or genetic inhibi- tion of the UFMylation system mitigates α -Syn secretion, whereas overexpression of UFBP1, a component of the endoplasmic reticulum–associated UFMylation ligase complex, augments α - 哺乳动物细胞和模型生物中的Syn分泌。UFM1本身与α -Syn共归因,而血清UFM1水平与α -Syn的水平相关。因为UFM1可以被泛素特异性肽酶19(USP19)直接识别,这是一种先前已建立的UCPS刺激剂,已知与多种伴侣活动相关,因此UFMylation可能会促进USP19的底物参与,从而允许对差异蛋白质进行严格选择,以使其对分泌蛋白进行严格的分泌和蛋白质的毒素有效性。

通过翻译后修改对胰岛素分泌的调节
胰岛素是一种从胰岛胰岛细胞中释放出的合成代谢激素,具有保持血糖稳态的独特能力(Cabrera等,2006; Da Silva Xavier,2018)。胰岛素对葡萄糖和其他营养素(例如氨基酸和游离脂肪酸)的分泌是一个复杂的过程,涉及多个信号通路的配位(Nolan等,2006; Henquin,2011)。这一过程的损害与糖尿病的发展直接相关(Schwartz等,2013)。因此,胰岛素分泌过程被认为是治疗糖尿病的有希望的靶标(Defronzo等,2014)。但是胰岛素分泌法规的细节仍然是一个未解决的问题。翻译后修饰(PTM)是在蛋白质上添加或去除化学基团的共价修改(Walsh等,2005)。通过调节蛋白质定位,降解和功能,它几乎与所有生理和病理过程密切相关(Walsh和Jefferis,2006; Khan等,2016; Morales-Tarre等,2021; Zhu and Hart and Hart,2021)。积累的证据表明,PTM广泛参与胰岛素分泌过程,目前,至少八种类型的PTMS已知与胰岛素分泌有关。例如,信号级联介导胰岛素分泌需要磷酸化(Campbell和Newgard,2021年)。sumoylation和棕榈酰化可以在多个阶段调节胰岛素分泌(Davey等,2019; Chamberlain等,2021)。这些表明乙酰化,泛素化和O-Glcnacylation与胰岛素基因转录有关(Mounier and Posner,2006; Ozcan等,2010; Sampley和Ozcan,2012)。甚至最近据报道,即使是一些研究的PTM,例如柠檬化和脱氨酸,也与胰岛素分泌有关。

大脑是通过分泌和副副转移的内分泌器官
这项研究报告说,副甲韧带(PTMS)被脑丘脑茎/祖细胞(HTNSC)分泌,以抑制受体细胞(例如纤维细胞)的衰老。释放后,PTM迅速转移到各种细胞类型的核中,包括神经元GT1-7细胞和不同的外围细胞,并有效地转移到体内各个大脑区域的神经元核中。值得注意的是,脑神经元还会产生和释放PTM,并且由于神经元种群很大,因此对于在脑脊液中维持PTM很重要,该PTM可以进一步转移到血液中。与其他几个大脑区域相比,下丘脑对长距离PTMS转移更强,支持该功能中关键的下丘脑作用。在生理学中,衰老与PTMS的产生和大脑转移的下降有关,并且研究了低丘脑与海马的PTMS敲低,显示出对神经行为生理学的不同贡献。总而言之,大脑是通过PTM的分泌和核转移的内分泌器官,以及下丘脑 - 该功能的大脑编排在生理学中具有保护性,并且针对衰老相关的疾病具有抵消性。
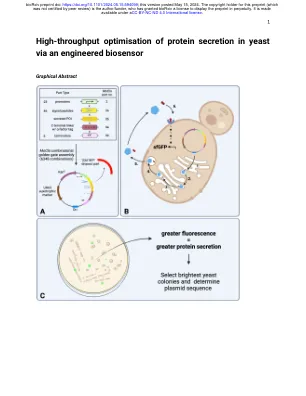
通过工程生物传感器
高价值蛋白质和酶的分泌是合成生物学经济的基础;允许在生产过程中连续发酵和蛋白质纯化而无需细胞裂解。大多数真核蛋白分泌由N末端信号肽编码;但是,信号肽序列变化对给定蛋白的分泌效率的强大影响尚未很好地定义。尽管天然信号肽序列多样性高,但大多数重组蛋白分泌系统仅采用少数表征良好的信号肽。此外,启动子和终止剂的选择可以显着影响分泌效率,但筛选众多遗传构建体以使最佳序列效率低下。在这里,我们调整了酵母G蛋白偶联受体生物传感器,以测量与任何感兴趣蛋白共归因于肽的肽标签的浓度。蛋白质分泌效率可以通过诱导受体激活下游的荧光报告基因来量化。此功能可以使用一锅组合金门克隆组装,对6000多个启动子,信号肽和终结器的6000多个组合进行高通量筛选。我们证明了这种生物传感器可以快速识别分泌和量化分泌水平的最佳组合。

铜绿假单胞菌型III分泌系统对免疫反应的调节
铜绿假单胞菌是一种革兰氏阴性细菌,引起免疫功能低下个体的感染。该病原体是Eskape病原体之一(包括粪肠球菌,金黄色葡萄球菌,克雷伯氏菌肺炎,baumanii,p.eruginosa,p.eruginosa,肠杆菌,肠杆菌,肠杆菌。),构成威胁生命的医院细菌(Hirsch和Tam,2010; Mulani等,2019)。铜绿假单胞菌还感染患有特定病理的患者,例如囊性纤维化(CF)。由于其形成生物膜的能力,铜绿假单胞菌通常会长期感染CF患者,并代表该疾病的负面结果(Malhotra等,2019)。为了成功地在宿主中建立自己,铜绿假单胞菌部署了一系列毒力因子,包括毒素,铁载体,粘附素和分泌系统(请参阅GONCgonçAlves-alves-de-albuquerque等人的评论,2016; Qin等,2022)。后者允许运输

评估电池租赁和销售电动车队的经济和环境影响:一项有关客户和公司含义的研究
β细胞功能障碍是糖尿病患者疾病进展的标志。研究一直集中在糖尿病发育过程中维持和恢复β细胞功能。这项研究的目的是探索人类胰岛中含有11A(Clec11a)的C型凝集素结构域的表达(CLEC11A),一种分泌的硫酸糖蛋白,并评估Clec11a对β细胞功能和体外增殖的影响。在这项研究中使用了这些假设,人类胰岛和人类βH1细胞系。我们确定了Clec11a在人类胰岛中的β细胞和α细胞中表达,但在内oc-βH1细胞中却没有表达,而在人类胰岛和Endoc-βH1细胞中都发现了CLEC11a的受体称为整合素亚基α11。用外源重组人Clec11a(RHCLEC11A)的长期治疗强调了葡萄糖刺激的胰岛素分泌,胰岛素含量以及来自人类胰岛和内c- H1细胞的增殖,这部分是由于转录因子MAFA和PDX1的强调表达水平。然而,在慢性棕榈酸酯暴露引起的β-βH1细胞中INS和MAFA的mRNA表达降低,只能通过引入RHCLEC11A来部分改善。基于这些结果,我们得出结论,RHCLEC11A促进了人β细胞中胰岛素的分泌,胰岛素含量和增殖,这与转录因子MAFA和PDX1的强调表达水平相关。clec11a可能会为糖尿病患者维持β细胞功能提供新的治疗靶点。

从大脑到生物标志物发现脑脊液的解密蛋白质分泌
摘要:脑脊液(CSF)是发现神经系统疾病生物标志物的重要基质。然而,CSF中蛋白质浓度的高动态范围阻碍了不靶向的质谱法检测最少丰富的蛋白质生物标志物。因此,对大脑内部的分泌过程有更深入的了解是有益的。在这里,我们旨在探讨脑蛋白是否以及如何预测CSF的分泌。通过将策划的CSF蛋白质组和人蛋白质图集的脑升高蛋白质组相结合,将脑蛋白分类为CSF或非CSF分泌。机器学习模型接受了一系列基于序列的特征的培训,以区分CSF和非CSF组,并有效地预测蛋白质的大脑起源。分类模型如果使用高置信度CSF蛋白,则在曲线下达到0.89的面积。最重要的预测特征包括亚细胞定位,信号肽和跨膜区域。分类器良好地概括为较大的大脑检测到的蛋白质组,并能够正确预测通过亲和力蛋白质组学鉴定的新型CSF蛋白。除了阐明蛋白质分泌的潜在机制外,受过训练的分类模型还可以支持生物标志物候选者的选择。关键字:脑蛋白质组,脑脊液,流体生物标志物,机器学习,蛋白质分泌■简介

脂肪细胞PI3K通过脂肪毒素效应
未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。此预印本的版权持有人(此版本发布于2023年4月26日。; https://doi.org/10.1101/2023.04.24.24.538076 doi:biorxiv Preprint