XiaoMi-AI文件搜索系统
World File Search Systemsynuclein

帕金森病的免疫系统
帕金森病的神经病理学特征是中脑腹侧多巴胺能神经元的退化、α-突触核蛋白 (α-syn) 聚集体在神经元中的积聚以及慢性神经炎症。在过去的二十年中,体外、离体和体内研究一致表明,小胶质细胞和星形胶质细胞介导的炎症反应参与其中,这些反应可能是由病理性 α-syn 或受影响的神经元和其他细胞类型的信号引起的,并且与神经退化和疾病发展直接相关。除了在中枢神经系统看到的显著免疫改变(包括 T 细胞向脑部的浸润)之外,最近的研究表明,先天和适应性区室的外周免疫特征发生了重要变化,尤其是单核细胞、CD4 + 和 CD8 + T 细胞。本综述旨在整合对帕金森病发病机制背后的免疫相关过程的综合理解,重点关注中枢和外周免疫细胞、神经元-神经胶质细胞串扰以及帕金森病发展过程中的中枢-外周免疫相互作用。我们的分析旨在全面了解帕金森病免疫机制的新知识及其对更好地理解该疾病整体发病机制的影响。

帕金森氏病中多巴胺能变性和中性粒细胞与淋巴细胞比的关联分析
关于PD的神经病理学特征,在SNPC中观察到了聚集的错误折叠的α-突触核蛋白夹杂物,通常称为Lewy身体,以及炎症的迹象(Moore等,2005)。最近的证据表明,神经炎症和周围炎症在PD的发作和进展中起着重要作用。关于神经炎症,McGeer的开创性研究最初建立了神经炎症与PD之间的联系。他观察到从PD患者获得的后大脑中激活的小胶质细胞浸润(McGeer等,1988)。在人类转录组学的荟萃分析中,Noori等。还表明,神经炎症是各种神经退行性疾病(例如路易斯体疾病(LBD)和其他非典型帕金森氏症)中的共同特征(Noori等人,2021年)。此外,来自动物模型和PD的验尸研究的证据表明,大脑受影响区域中活化的小胶质细胞和星形胶质细胞的过度表达异常,主要是在SNPC中(Gu等,2010; Colonna; Colonna; Colonna and Butovsky,2017)。此外,越来越有令人信服的证据表明,周围炎症与PD的早期病理生理有关,并且在整个疾病过程中都会发生动态变化(Pajares,2020年)。

通过体育锻炼增强突触核病的睡眠质量
在睡眠期间,发生了几个至关重要的脑稳态过程,包括突触连接的重排,这对于记忆形成和更新至关重要。睡眠还促进了神经毒性废物的去除,其积累在神经变性中起着关键作用。各种神经成分和环境因素调节和影响清醒和睡眠之间的生理转变。在这种复杂系统中的破坏构成了睡眠障碍的基础,如在突触核生物病中所观察到的那样。突触核酸是神经退行性疾病,其特征在于大脑中α-突触核蛋白蛋白聚集体的异常。在不同的大脑区域中积累会导致一系列临床表现,包括低因素,认知障碍,精神病症状和神经疗法障碍。睡眠障碍在突触核心腺病患者中非常普遍,它们不仅会影响患者的整体福祉,而且还会直接导致疾病的严重程度和进展。因此,制定有效的治疗策略以改善这些患者的睡眠质量至关重要。足够的睡眠对于大脑健康至关重要,必须考虑突触中的突触中的作用在破坏睡眠模式中的作用。在这种情况下,必须探索体育锻炼作为一种潜在的非药理学干预措施来管理突触核心腺病患者的睡眠障碍。讨论了有关锻炼计划提高该患者人群睡眠质量功效的当前证据。
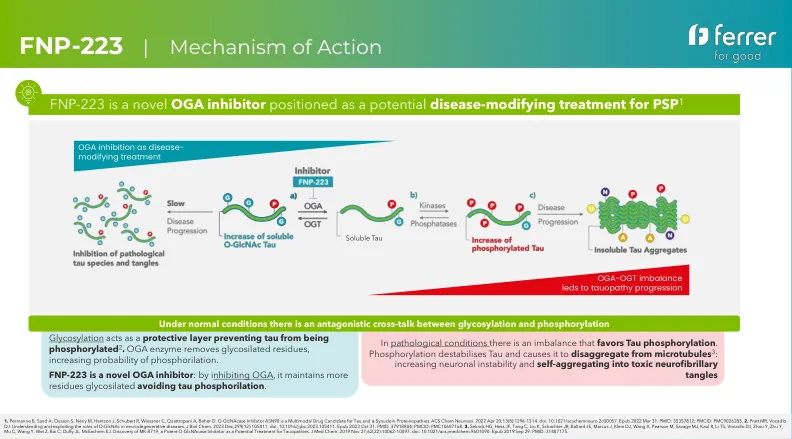
FNP-223 |作用机理
1。Permanne B,Sand A,Ousson S,NényM,Hantson J,Schubert R,Wiessner C,Quattropani A,Beher D. O -Glcnacase抑制剂ASN90是TAU和α-蛋白核蛋白蛋白质病的多型药物候选药物。ACS Chem Neurosci。2022 Apr 20; 13(8):1296-1314。 doi:10.1021/acschemneuro.2c00057。EPUB 2022 3月31日。PMID:35357812; PMCID:PMC9026285。 2。 Pratt MR,Vocadlo DJ。 理解和利用O-GLCNAC在神经退行性疾病中的作用。 J Biol Chem。 2023 DEC; 299(12):105411。 doi:10.1016/j.jbc.2023.105411。 EPUB 2023 10月31日。 PMID:37918804; PMCID:PMC10687168。 3。 Selnick HG,Hess JF,Tang C,Liu K,Schachter JB,Ballard JE,Marcus J,Klein DJ,Wang X,Pearson M,Savage MJ,Kaul R,Kaul R,Li TS,Vocadlo DJ,Zhou Y, 发现MK-8719,这是一种有效的O-Glcnacase抑制剂,是对功的潜在治疗方法。 J Med Chem。 2019年11月27日; 62(22):10062-10097。 doi:10.1021/acs.jmedchem.9b01090。 EPUB 2019年9月29日。 PMID:31487175。PMID:35357812; PMCID:PMC9026285。2。Pratt MR,Vocadlo DJ。理解和利用O-GLCNAC在神经退行性疾病中的作用。J Biol Chem。 2023 DEC; 299(12):105411。 doi:10.1016/j.jbc.2023.105411。 EPUB 2023 10月31日。 PMID:37918804; PMCID:PMC10687168。 3。 Selnick HG,Hess JF,Tang C,Liu K,Schachter JB,Ballard JE,Marcus J,Klein DJ,Wang X,Pearson M,Savage MJ,Kaul R,Kaul R,Li TS,Vocadlo DJ,Zhou Y, 发现MK-8719,这是一种有效的O-Glcnacase抑制剂,是对功的潜在治疗方法。 J Med Chem。 2019年11月27日; 62(22):10062-10097。 doi:10.1021/acs.jmedchem.9b01090。 EPUB 2019年9月29日。 PMID:31487175。J Biol Chem。2023 DEC; 299(12):105411。 doi:10.1016/j.jbc.2023.105411。EPUB 2023 10月31日。PMID:37918804; PMCID:PMC10687168。3。Selnick HG,Hess JF,Tang C,Liu K,Schachter JB,Ballard JE,Marcus J,Klein DJ,Wang X,Pearson M,Savage MJ,Kaul R,Kaul R,Li TS,Vocadlo DJ,Zhou Y,发现MK-8719,这是一种有效的O-Glcnacase抑制剂,是对功的潜在治疗方法。J Med Chem。 2019年11月27日; 62(22):10062-10097。 doi:10.1021/acs.jmedchem.9b01090。 EPUB 2019年9月29日。 PMID:31487175。J Med Chem。2019年11月27日; 62(22):10062-10097。 doi:10.1021/acs.jmedchem.9b01090。 EPUB 2019年9月29日。 PMID:31487175。2019年11月27日; 62(22):10062-10097。 doi:10.1021/acs.jmedchem.9b01090。EPUB 2019年9月29日。PMID:31487175。

昼夜节律基因BMAL1的神经元缺失诱导细胞自主多巴胺能神经变性
昼夜节律功能障碍是帕金森病(PD)的标志,在PD患者中已经描述了核心时钟基因BMAL1的表达降低。bmal1是核心昼夜节律函数所必需的,但也具有非节律函数。种系BMAL1缺失会导致小鼠的脑氧化应激和突触丧失,并且会加剧多巴胺能神经变性,以响应毒素MPTP。在这里,我们检查了细胞类型 - 特异性BMAL1缺失对体内多巴胺能神经元活力的影响。我们观察到,BMAL1的全球,产后缺失导致酪氨酸羟化酶 +(Th +)多巴胺能神经元的自发丧失。这不是通过光诱导的行为昼夜节律破坏来复制的,也不是由星形胶质细胞或小胶质细胞特异性BMAL1缺失引起的。然而,泛神经元或神经元特异性BMAL1缺失会导致SNPC中Th +神经元的细胞自主丧失。bmal1缺失并未改变α-突触核蛋白原纤维注射后神经元丧失的百分比,尽管BMAL1 -KO小鼠在基线时的神经元较少。转录组学分析表明,参与氧化磷酸化和帕金森氏病的途径失调。这些发现证明了BMAL1在调节多巴胺能神经元存活中的细胞自主作用,并且可能对PD的神经保护具有重要意义。

溶酶体存储,高彻和帕金森氏病的自噬和先天免疫受损:药物发现的见解
自噬 - 溶酶体途径的损害越来越涉及帕金森氏病(PD)。GBA1突变引起溶酶体储存障碍Gaucher病(GD),是PD的最常见遗传危险因素。GBA1突变已显示会引起自噬 - 溶酶体损伤。 不良细胞成分的自噬降解有缺陷与多种病理有关,包括正常蛋白质稳态的丧失,特别是α-突触核蛋白和先天免疫功能障碍。 在PD和GD中观察到后者。 在这里,我们将讨论自噬和免疫失调之间的机理联系,以及这些病理学在肠道和大脑之间在这些疾病中的沟通中的可能作用。 在神经性GD(NGD)的蝇模型中的最新工作显示肠自噬缺陷导致胃肠道功能障碍和免疫激活。 雷帕霉素治疗部分逆转了自噬阻滞并降低了免疫活性,与生存率增加并改善了运动能力。 肠道微生物组的改变是神经炎症的关键驱动力,研究表明,在NGD蝇中消除了微生物组,而PD的小鼠模型可以改善脑部炎症。 在这些观察结果之后,将溶酶体 - 自噬途径,先天免疫信号传导和微生物组营养不良症讨论为PD和GD中的潜在治疗靶标。 本文是讨论会议问题的一部分,“理解神经变性中的内聚糖网络”。GBA1突变已显示会引起自噬 - 溶酶体损伤。不良细胞成分的自噬降解有缺陷与多种病理有关,包括正常蛋白质稳态的丧失,特别是α-突触核蛋白和先天免疫功能障碍。在PD和GD中观察到后者。在这里,我们将讨论自噬和免疫失调之间的机理联系,以及这些病理学在肠道和大脑之间在这些疾病中的沟通中的可能作用。在神经性GD(NGD)的蝇模型中的最新工作显示肠自噬缺陷导致胃肠道功能障碍和免疫激活。雷帕霉素治疗部分逆转了自噬阻滞并降低了免疫活性,与生存率增加并改善了运动能力。肠道微生物组的改变是神经炎症的关键驱动力,研究表明,在NGD蝇中消除了微生物组,而PD的小鼠模型可以改善脑部炎症。在这些观察结果之后,将溶酶体 - 自噬途径,先天免疫信号传导和微生物组营养不良症讨论为PD和GD中的潜在治疗靶标。本文是讨论会议问题的一部分,“理解神经变性中的内聚糖网络”。

来自细胞系的多组学和药物筛选数据的统计整合
数据集成方法用于获得多个数据集的统一摘要。对于多模态数据,我们提出了一种计算工作流程来联合分析来自细胞系的数据集。该工作流程包括一种用于多组学数据的新型概率数据集成方法,称为 POPLS-DA。该工作流程的动机是对突触核蛋白病的研究,其中在受影响的 LUHMES 细胞系和对照中测量转录组学、蛋白质组学和药物筛选数据。目的是突出显示与突触核蛋白病有关的潜在可用药途径和基因。首先,使用 POPLS-DA 优先考虑最能区分病例和对照的基因和蛋白质。对于这些基因,构建了一个集成的相互作用网络,其中结合了药物筛选数据以突出显示网络中的可用药基因和途径。最后,进行功能富集分析以识别保护性药物靶向的突触和溶酶体相关基因和蛋白质簇。将 POPLS-DA 与其他单组学和多组学方法进行了比较。我们发现,热休克蛋白 70 家族成员 HSPA5 是经过验证的药物(尤其是 AT1 阻滞剂)最常针对的基因之一。HSPA5 和 AT1 阻滞剂之前已被证实与 α-突触核蛋白病理和帕金森病有关,这表明我们的发现具有相关性。我们的计算工作流程确定了治疗突触核蛋白病的新方向。与其他单组学和多组学方法相比,POPLS-DA 提供了更大的可解释基因集。基于 R 和 markdown 的实现可在线免费获取。

亚细胞结构对Rotenone在SH-SY5Y细胞中诱导的发病机理的脆弱性
神经退行性疾病可以被识别为一类神经疾病,在细胞内病理学过程中共享相似性。神经变性的最典型标志,特异性蛋白的积累,伴随着其他细胞内过程,导致亚细胞结构功能障碍。在帕金森氏病,α-突触核蛋白(αs)的积累和高级聚集体的形成中,最终被认为是Lewy体的,被认为是主要的标志(Braak等人2003)。 帕金森氏病(PD)是全球第二普遍的神经退行性疾病(Polito等人 2016)。 在PD病理学期间发生了多种平行和/或先前的变化。 鉴于有关该主题的数据量,线粒体功能障碍主要是通过呼吸链破坏并因此增加了活性氧(ROS)的产生,仍被认为在细胞内发病机理中起着不可或缺的作用(Perfeito等人(Perfeito等) 2012,Park等。 2018)。 线粒体网络通过平衡的融合和裂变过程积极维持,通过线粒体来协调线粒体降解(Park等人 ) 2018)。 线粒体膜动力学受分子因子范围适当调节。 除其他外,αs主要由神经元细胞表达,并且似乎与线粒体动态过程密切相关。 αs的过表达对线粒体生理具有毒性作用。 更高2003)。帕金森氏病(PD)是全球第二普遍的神经退行性疾病(Polito等人2016)。在PD病理学期间发生了多种平行和/或先前的变化。鉴于有关该主题的数据量,线粒体功能障碍主要是通过呼吸链破坏并因此增加了活性氧(ROS)的产生,仍被认为在细胞内发病机理中起着不可或缺的作用(Perfeito等人(Perfeito等)2012,Park等。 2018)。 线粒体网络通过平衡的融合和裂变过程积极维持,通过线粒体来协调线粒体降解(Park等人2012,Park等。2018)。线粒体网络通过平衡的融合和裂变过程积极维持,通过线粒体来协调线粒体降解(Park等人2018)。线粒体膜动力学受分子因子范围适当调节。除其他外,αs主要由神经元细胞表达,并且似乎与线粒体动态过程密切相关。αs的过表达对线粒体生理具有毒性作用。 更高αs的过表达对线粒体生理具有毒性作用。更高

帕金森病的深部脑刺激机制
帕金森病 (PD) 是第二大最常见的神经退行性疾病,其发病率随着年龄增长而上升,男性更容易患上该病 [1]。目前,PD 缺乏确切的诊断方法,因此临床诊断仍然是确诊的基本依据 [2,3]。医护人员根据主要症状进行临床诊断,并使用诊断标准排除其他可能的原因 [2,3]。PD 的典型运动症状包括静止性震颤、运动迟缓和僵硬 [4]。根据研究和统计方法的不同,估计全球每年 PD 发病率在十万人中 8.7 至 19 人之间 [5]。目前,全球 PD 患者超过 1000 万 [6]。研究人员普遍认为 PD 是一种受多种因素影响的复杂疾病。这些因素包括遗传因素(常染色体显性、常染色体隐性、易感基因)、环境因素(如接触碳氢化合物)、便秘、体力活动、吸烟(尼古丁)和咖啡因摄入量 [7-9]。该疾病被认为是由于黑质 (SN) 最初受损,特别是其致密区受损,导致 SN 活动减弱,同时基底神经节其他区域 [包括丘脑底核 (STN)] 抑制丧失,从而引起过度活动 [10]。PD 的典型病理变化包括中脑多巴胺能 (DAergic) 神经元的快速丢失以及脑内 α - 突触核蛋白聚集体形成的路易体数十年的积累 [9,11,12]。

代谢能量下降的偶联性失调在生理上高度活跃的神经元中的儿茶酚胺代谢:对帕金森氏病选择性神经元死亡的影响
帕金森氏病(PD)是一种与年龄相关的不可逆性神经退行性疾病,其特征在于,由于nigra nigra pars pars compacta(SNPC)的多巴胺能(DA)神经元的丧失引起的一种逐渐恶化的非自愿运动障碍。PD的两个主要病理生理特征是受影响神经元中包含体的积累,以及在Nigra pars compacta(SNPC)(SNPC)和氯肾上腺素(LC)中含有神经元素的DA神经元的主要丧失。包含体包含错误折叠和聚集的α-核蛋白(α -syn)纤维,称为刘易体。PD的病因和致病机制是复杂的,多维的,并且与环境,遗传和其他与年龄有关的因素的组合相关。尽管已经广泛研究了与PD的致病机制相关的个体因素,但尚未设想发现发现与统一的致病机制的整合。在这里,我们提出了一种基于当前可用的实验数据的独特的高代谢活性耦合的高代谢活性耦合的升高能量需求,提出了PD中SNPC和NE神经元变性的综合机制。所提出的假设机制主要基于这些神经元的独特高代谢活性升高的升高。我们认为,在PD中,SNPC和NE神经元中选择性的DA神经元的高脆弱性可能是由于细胞能量调节。这种细胞能量调节可能会引起这些神经元中氧化还原活性金属稳态(尤其是铜和铁)的DA和NE代谢失调。