XiaoMi-AI文件搜索系统
World File Search System分选

通过工程改造的人类 Treg 稳定表达 FOXP3 和核心耐受性基因
FOXP3+ 调节性 T 细胞 (Tregs) 在预防致命自身免疫和维持组织稳态方面发挥着关键作用。Tregs 的功能稳定性对于其对免疫耐受而非失控免疫的贡献仍然至关重要,特别是对于细胞疗法而言,炎症微环境可能会影响 FOXP3 和相关耐受性基因的表达。为了解决潜在的 Treg 不稳定性,我们通过 PBMC 分离的 CD4+ T 细胞的基因编辑方法生成了人类工程化 Tregs (EngTregs),从而导致稳定的 FOXP3 表达和雷帕霉素激活信号复合物,可提供可调的 IL-2 信号,从而有效地将 FOXP3 表达与已知在炎症条件下促进 Treg 不稳定的现有调节元件分离。使用转录组分析,我们评估了培养的分选 Tregs (cTregs) 与 EngTregs 相比的稳定性。基于大量和单细胞 RNA 测序分析,我们发现 EngTregs 表达的关键“核心 Treg”和“FOXP3 协同”基因水平更高,这通过流式细胞术得到证实。此外,与 cTregs 相比,在 EngTregs 中观察到关键 Treg 稳定性标志物 CD27、CD70 和 IKZF4 (EOS) 的有利表达模式。相反,与 EngTregs 相比,cTregs 表达更高水平的细胞毒性基因,包括 GZMA 和 PERFORIN1 以及其他炎症基因。最后,我们观察到这些相关性具有功能意义,这通过与 cTregs 相比,EngTregs 中关键耐受性标志物的表达更高来证明。这项研究有力地支持了 EngTregs 是稳定的、耐受性的 FOXP3+ T 细胞,从而为治疗危及生命的自身免疫和自身炎症疾病提供了宝贵的资产。
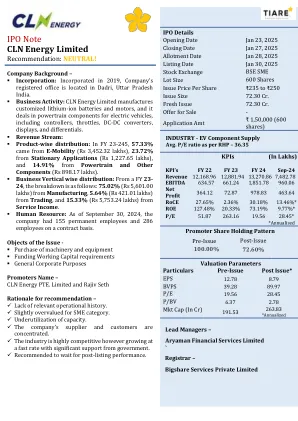
IPO注意CLN Energy Limited
该公司计划在其NOIDA设施中扩大制造业,将两轮电池测试能力提高69.4%(从72 MWH到122 MWH)。对于其他电池,容量将显着上升:细胞分级为348.8%(41至184 MWH),细胞分选49.1%(110至164 MWH),制造43.5%(115至165 MWH),并且电池测试量为243.9%(41至141 MWH)。这项战略投资可增强产量,提高效率并增强市场竞争力,同时支持不断增长的需求和运营可伸缩性。该公司旨在扩大其产品组合,以满足对锂离子电池和动力总成组件的不断增长,这是由EV Boom和该国向绿色革命转移的驱动的。在FY23-24(直到2024年9月)中,电池占收入的82.34%(4,958.27千万卢比),电动机和控制器10.67%(642.73千万卢比),收款2.53%(152.50千万卢比),其他4.46%(4.46%)(4.46%)(268.52 CRORES)。 该公司试图确定电池的新用例,并利用未利用的能力,以增强其在国内市场的可见性,增加客户对锂离子电池的支出,并增强其在发电和存储段中的地位。在FY23-24(直到2024年9月)中,电池占收入的82.34%(4,958.27千万卢比),电动机和控制器10.67%(642.73千万卢比),收款2.53%(152.50千万卢比),其他4.46%(4.46%)(4.46%)(268.52 CRORES)。该公司试图确定电池的新用例,并利用未利用的能力,以增强其在国内市场的可见性,增加客户对锂离子电池的支出,并增强其在发电和存储段中的地位。

阿莫西林 - 耐肺炎链球菌可以是...
重要的机会性人类病原体肺炎链球菌中的抽象抗生素抗性正在上升。在β-乳酰胺抗生素阿莫西林(这是一线疗法)的情况下,这尤其有问题。因此,发现杀死或对抗阿莫西林耐药性肺炎球菌的靶标至关重要。为此,我们使用称为Scrilecs-Seq的CRISPR干扰(CRISPR干扰库的亚集)开发了一个全基因组,基于单细胞的基因沉默屏幕,该筛选是由荧光激活的细胞分选提取的,耦合与下一代测序)。由于阿莫西林会影响生长和分裂,因此使用SCRILECS-SEQ来识别负责维持适当细胞大小的靶标。我们的屏幕表明,大甲酸酯途径的下调会导致广泛的细胞伸长。进一步研究这种现象,这表明它是由于细胞壁合成部位在细胞壁合成部位的可用性降低而引起的,这是由于未依赖磷酸盐(UND-P)的限制,这是脂质载体,该脂质载体负责将这些前体跨细胞膜运输。数据表明,即使肽聚糖的合成仍在继续,即使降低了UND-P水平,但细胞收缩也被专门停止。我们成功利用了这一知识,以创建一种组合治疗策略,其中FDA批准的药物氯米芬是一种UND-P合成的抑制剂,与阿莫西林配对。我们的结果表明,克罗米芬增强了阿莫西林蛋白的抗菌活性,并且联合疗法使耐肺炎链球菌恢复活力。这些发现可以提供一个起点,以开发越来越多的难以治疗的抗肺炎球菌感染的解决方案。

一种增强荚膜红细菌产氢的定向基因组进化方法
光合细菌(如红细菌)的固氮酶依赖性 H 2 生成已被广泛研究。使用基因操作增加 H 2 产量的一个重要限制是缺乏高通量筛选方法来检测可能的过量生产突变体。之前,我们设计了红细菌菌株,使其在 H 2 反应中发出荧光,并利用它们来识别导致 H 2 过量生产的固氮酶 Fe 蛋白突变。在这里,我们使用紫外线在工程 H 2 感应菌株的基因组中诱导随机突变,并使用荧光激活细胞分选从含有 5 × 10 5 突变体的文库中检测和分离 H 2 过量生产细胞。三轮诱变和菌株选择逐渐使 H 2 产量增加了 3 倍。对五种 H 2 过量生产菌株的全基因组进行了测序,并与亲本感应菌株的全基因组进行了比较,以确定 H 2 过量生产的基础。除了转录激活因子 nifA2 之外,与氮固定相关的已知功能没有发生突变。然而,一些突变被映射到能量产生系统和碳代谢相关功能,这些功能可以将还原力或 ATP 提供给固氮酶。在批量培养中,固氮酶抑制的时间过程实验揭示了固氮酶蛋白水平与其 H 2 和乙烯生产活动之间的不匹配,这表明能量受到限制。在恒化器中培养产生的 H 2 始终比相应的批量培养多 19 倍,揭示了选定的 H 2 过量生产菌株的潜力。

肽耦合的RBC作为自身免疫性疾病的治疗
a Anti AAV Adeno-Associated Virus acc Acceleration ANOVA Analysis of Variance APC Antigen Presenting Cells APL Altered peptide ligands AUC Area Under the Curve BBB Blood-Brain Barrier Blvrb Biliverdin Reductase b BMDC Bone Marrow Derived Dendritic Cells BMDM Bone Marrow Derived Macrophages BTLA B And T Lymphocyte Associated CCL Chemokine (C-C motif) Ligand CCR C-C Chemokine Receptor CD Cluster of Differentiation Cdh5 Cadherin 5 CFA Complete Freund's Adjuvant CFSE Carboxyfluorescein Succinimidyl Ester CIS Clinically Isolated Syndrome CLEC4F C-type lectin domain family 4 member F CNS Central Nervous System CSF Cerebrospinal Fluid CTFR Cell Trace Far Red CTLA-4 Cytotoxic T-lymphocyte-Associated Protein 4 DCs Dendritic Cells dec Deceleration DEGs Differentially Expressed Genes DMT Disease-modifying Therapies DTx Diptheria Toxin EAE Experimental Autoimmune Encephalomyelitis EBV Epstein-Barr Virus EDC 1-Ethyl-3-(3-dimethylaminopropyl) Carbodiimide EDSS扩展的残疾状态量表E FACS荧光细胞分选FBS胎牛血清FCGR FCGR FC FC FC FC FC FC受体FCNA FICOLIN 1 FDR FRASE FALSE发现率FGCZ功能基因组中心Zurich Foctimation Center Zurich Focp3 Foxp3 Forkhead Forkhead Box蛋白3 GDF15生长/分化因子15 gdf15 gdf15 gdf15 gdf15

双链到单链的转变在 DNA 折纸纳米结构中引起力和运动
除静态纳米结构外,DNA纳米技术还能构建动态和自主开关。[18] 这些动态开关的操作可分为两大类:第一,通过分子相互作用操作;第二,通过外部刺激操作。用于控制纳米尺度运动的主要分子相互作用是DNA杂交(主要是立足点介导的链置换)和碱基堆积。由分子相互作用控制的此类运动的例子包括可重构等离子体装置、[19] 铰链、[20,21] 镊子、[18,22] 旋转装置、[23–26] 助行器、[27] 药物载体 [28,29] 和对分子或纳米颗粒进行分选的机器人。[30,31] 作为驱动机制的其他分子相互作用包括靶分子结合 [32,33] 和适体 [28,29] 以及核小体相互作用。 [34] 通过任何分子相互作用进行的操作(包括上述所有机制)具有可控分子识别和特异性的优点。 然而,操作速度受到分子扩散和相互作用动力学的限制,因此通常非常慢。 值得注意的是,已经开发出多种方法来提高动态 DNA 装置的响应速度。 另一方面,外部刺激如光、[35,36] 温度、[37] 离子、[11,23] pH、[38–40] 和电场 [21,41] 通常能够使操作速度提高很多个数量级。[41] 例如,Karna 等人利用相邻纳米结构域之间可逆的、pH 依赖性的 i-基序形成来促进卷曲 DNA 纳米弹簧的驱动,进而通过整合素偶联影响培养细胞的运动性。 [40] 然而,我们在此称之为外部刺激的任何一种,都存在着整体作用的局限性,而且缺乏分子识别所能提供的特异性。

密码子
抽象背景:黑色素瘤是皮肤癌最具侵略性的形式。黑色素瘤干细胞(MSC)是黑色素瘤侵袭和转移的驱动力之一。因此,探索维持MSC茎的机制非常重要。在这项研究中,表征了从A375细胞系衍生的CD147阳性(CD147+)MSC。方法:从A375细胞中对侧种群(SP)和非SP细胞进行分选。进行了定量的实时聚合酶链反应和蛋白质印迹分析,以确定SP和非SP细胞中CD147的表达。随后,从SP细胞中分离出CD147+和CD147阴性(CD147-)细胞。通过球体形成,伤口 - 修复和Transwell分析,可以鉴定出CD147 +/-抗原呈现细胞的干细胞特征和转移潜力。Western印迹分析,以评估反式形成生长因子-BETA1(TGFβ1)和神经源性基因座缺口同源蛋白1(Notch1)信号通路的蛋白质水平。异种移植肿瘤实验,以研究体内CD147+细胞的肿瘤基因能力。结果:CD147在A375细胞系的SP细胞中高度表达。CD147+细胞在体外具有更强的球体形成,迁移和侵袭的能力。CD147+细胞中TGFβ1,Notch1,Jagged1和Hes1的蛋白质水平高于CD147-细胞。此外,CD147+细胞在体内显示出更强的致瘤和转移性潜力。©2024密码子出版物。由密码子出版。结论:A375细胞系的SP细胞表达了高水平的CD147,而CD147+ SP细胞POS具有更强的茎样特征和运动性,这与TGFβ和Notch途径的激活有关。

新鲜的冷冻血浆可能会降低内皮水平
背景:重大烧伤的患者与结晶和胶体的组合复苏。在烧伤复苏期间,还可以将新鲜的冷冻血浆(FFP)作为辅助性胶体溶液燃烧的患者(TBSA)燃烧。FFP可能会减少与大型TBSA烧伤相关的内皮功能障碍。此外,FFP可能通过影响脂肪衍生的干细胞释放的细胞因子水平(ADSC),特别是细胞因子VEGFF-A来改变患者的炎症状态。这项研究旨在研究FFP对烧伤患者VEGF-A水平的作用。方法:在IRB批准后,在初次手术期间从成年患者中收集脂肪组织。ADSC。荧光激活的ADSC的单细胞分选(FACS),以确定CD105,CD90和CD73抗体的纯度。ADSC在标准组织培养条件下生长,并收集上清液进行细胞因子分析。使用线性回归分析数据,以将FFP的总量与VEGF-A水平绘制总量以及Spearman相关性。结果:这项研究纳入了燃烧后36小时内接受FFP的14名患者。给出的FFP量为258-3186毫升,平均为1465±715 ml。这些患者的平均TBSA为42±22%,平均患者年龄为53±16岁。未来的研究需要增加样本量以支持这一发现。线性回归和Spearman相关性均显示出FFP量之间的中等强度相关性(r = -0.5758和Spearman系数= -0.433)。结论:VEGF-A先前已被证明在血管生成中起作用,这可以增加炎症细胞浸润并导致内皮细胞功能障碍。接受较高FFP水平的患者与较低水平的VEGF-A相关,表明较高剂量的FFP和降低的内皮细胞功能障碍之间可能存在相关性。

RecT 重组酶表达可实现高效的基因...
摘要 屎肠球菌是一种普遍存在的革兰氏阳性细菌,可从环境、食物和哺乳动物的微生物群中分离得到。屎肠球菌的共生菌株可对宿主的生理和免疫产生有益影响,但抗生素的使用已从牲畜和人类中产生了抗生素耐药性和致病性分离株。然而,由于基因编辑方法效率低下,对屎肠球菌功能和机制的解析受到限制。为了解决这些限制,我们在此报告,屎肠球菌 RecT 重组酶的表达可显著提高重组工程技术在屎肠球菌的共生菌株和抗生素耐药菌株以及其他肠球菌种(如 E. durans 和 E. hirae)中的效率。值得注意的是,RecT 与成簇的规律间隔短回文重复序列 (CRISPR)-Cas9 和向导 RNA (gRNA) 的结合表达能够实现高效的无瘢痕单链 DNA 重组,从而在屎肠球菌中产生特定的基因编辑突变体。此外,我们证明屎肠球菌 RecT 表达促进了编码抗生素选择标记的双链 DNA 模板的染色体插入,从而产生了基因缺失突变体。作为进一步的原理证明,我们使用 CRISPR-Cas9 介导的重组敲除屎肠球菌中的两个分选酶 A 基因,以进行下游功能表征。此处描述的通用 RecT 介导的重组方法应能显著增强对屎肠球菌和其他密切相关物种的遗传研究,以进行功能和机制研究。

对“小胶质激活状态驱动神经退行性疾病的葡萄糖摄取和FDG-PET改变”的评论的回应
引起了极大的兴趣,我们认识到Zimmer等人的技术评论。Xiang等人在我们最近的科学转化医学论文中。 (1)。 我们感谢关于氟脱氧葡萄糖正电子发射断层扫描(FDG-PET)信号的细胞起源的讨论,并且考虑到剩下的许多开放问题,我们希望这将刺激该领域的进一步重要研究。 我们完全同意Zimmer等人。 星形胶质细胞在大脑的FDG摄取中起着重要作用,并且我们没有否认星形胶质细胞FDG吸收的贡献是整体FDG-PET信号的主要来源。 但是,考虑到另一组(2)的最新数据,以及我们的发现(1),小胶质细胞FDG摄取对FDG-PET信号的实质性直接贡献,特别是在髓样细胞2(trem2)表达的Trig Gering受体2(trem2) - 依赖性激活的激活非常重要的情况下,重要的是要考虑。 首先,在技术评论中,Zimmer等人。 表明,总小胶质细胞数量可能太低,无法影响总FDG-PET信号。 但是,我们的数据清楚地表明,小胶质细胞是淀粉样小鼠模型中FDG-PET的增加,并且在TREM2-KNOCKOUT(TREM2-KO)小鼠中降低了小胶质细胞,而小胶质细胞占健康小鼠脑中FDG总吸收的约10%。 小胶质细胞对FDG摄取的贡献是由野生型小鼠的99%小胶质细胞耗竭后的PET信号降低的幅度确定的。 我们在分离细胞中绝对活性的交叉校准后重新评估了我们的数据,并在分离前整个大脑的活性。 1,a和b)(3)。 1C)。Xiang等人在我们最近的科学转化医学论文中。(1)。我们感谢关于氟脱氧葡萄糖正电子发射断层扫描(FDG-PET)信号的细胞起源的讨论,并且考虑到剩下的许多开放问题,我们希望这将刺激该领域的进一步重要研究。我们完全同意Zimmer等人。星形胶质细胞在大脑的FDG摄取中起着重要作用,并且我们没有否认星形胶质细胞FDG吸收的贡献是整体FDG-PET信号的主要来源。但是,考虑到另一组(2)的最新数据,以及我们的发现(1),小胶质细胞FDG摄取对FDG-PET信号的实质性直接贡献,特别是在髓样细胞2(trem2)表达的Trig Gering受体2(trem2) - 依赖性激活的激活非常重要的情况下,重要的是要考虑。首先,在技术评论中,Zimmer等人。表明,总小胶质细胞数量可能太低,无法影响总FDG-PET信号。但是,我们的数据清楚地表明,小胶质细胞是淀粉样小鼠模型中FDG-PET的增加,并且在TREM2-KNOCKOUT(TREM2-KO)小鼠中降低了小胶质细胞,而小胶质细胞占健康小鼠脑中FDG总吸收的约10%。小胶质细胞对FDG摄取的贡献是由野生型小鼠的99%小胶质细胞耗竭后的PET信号降低的幅度确定的。我们在分离细胞中绝对活性的交叉校准后重新评估了我们的数据,并在分离前整个大脑的活性。1,a和b)(3)。1C)。我们现在提供了一个额外的FDG分配模型,该模型考虑了示踪剂注射后通过磁性激活的细胞分选评估的单胶质细胞摄取(1),考虑到小鼠大脑中的7%小胶质细胞(图在这里,我们发现了对小胶质细胞对PET(99%耗竭时降低9.8%)和磁性细胞分选的相对贡献的相对贡献(MAC; 8.6%;图。还观察到淀粉样小鼠模型的小胶质细胞对PET(29.1%)和MAC(29.1%)和MAC(29.9%)的总FDG贡献之间的良好一致性,假设由于繁殖而导致的小胶质细胞密度增加了1.3倍(1)。