XiaoMi-AI文件搜索系统
World File Search System合成酶

定向进化 Methanomethylophilus alvus 吡咯氨酰-tRNA 合成酶可产生高活性和高选择性变体
吡咯赖氨酸-tRNA 合成酶(PylRS)通常用于将非规范氨基酸(ncAA)位点特异性掺入蛋白质中。最近,Methanomethylophilus alvus PylRS(Ma PylRS)的活性位点经过合理设计,以扩大其底物兼容性,从而能够掺入难以结合的 ncAA。然而,尚未报道活性位点以外的可增强 Ma PylRS 酶特性的突变。我们利用噬菌体辅助非连续进化(PANCE)来进化 Ma PylRS,以有效掺入 N ε -Boc- L -赖氨酸(BocK)。定向进化产生了活性位点外的几种突变,这些突变大大提高了酶的活性。我们结合最有效的突变来生成一种新的 PylRS 变体(PylRS opt),它对几种赖氨酸和苯丙氨酸衍生物具有高活性和选择性。 PylRS opt 中的突变可用于增强先前设计的 PylRS 构建体,例如 Ma PylRS N166S,并且 PylRS opt 适用于需要双 ncAA 掺入的应用,并可显著提高这些目标蛋白的产量。

通过化学抑制 4′-磷酸泛酰-l-半胱氨酸合成酶 (CoaB) 活性来靶向结核分枝杆菌 CoaBC
摘要:辅酶 A (CoA) 是所有活细胞中普遍存在的辅助因子,据估计多达 9% 的细胞内酶促反应都需要它。结核分枝杆菌 (Mtb) 依靠自身生物合成 CoA 的能力来满足依赖这种辅因子发挥活性的无数酶促反应的需要。因此,CoA 生物合成途径被认为是新型结核病药物靶点的潜在来源。在之前的工作中,我们在体内和体外通过基因验证了 CoaBC 是 Mtb 的杀菌药物靶点。在这里,我们描述了化合物 1f 的鉴定,它是双功能 Mtb CoaBC 的 4′-磷酸泛酰-L-半胱氨酸合成酶 (PPCS;CoaB) 结构域的小分子抑制剂,并表明该化合物在 Mtb 中表现出靶向活性。发现化合物 1f 对 CoaBC 的抑制作用与 4 ' - 磷酸泛酸(CoaB 催化反应的底物)不具竞争性。此外,野生型 Mtb H37Rv 在暴露于化合物 1f 后进行的代谢组学分析产生了与泛酸和 CoA 生物合成扰动一致的特征。作为首次报道的 Mtb CoaBC 直接小分子抑制剂,该抑制剂具有靶向选择性全细胞活性,本研究证实了 CoaBC 的药物可行性,并从化学上验证了该靶点。关键词:结核病、药物发现、辅酶 A、CoaBC
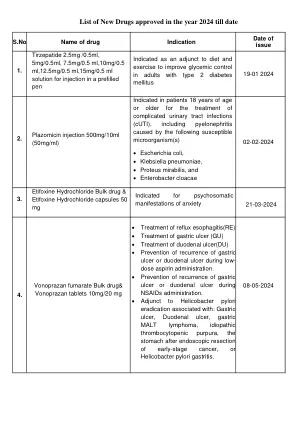
在2024年批准的新药清单
在尿素周期疾病的慢性治疗中表示为辅助治疗,涉及磷酸氨基磷酸氨基磷酸盐合成酶(CPS),鸟氨酸经钙化酶(OTC)或精氨酸糖酸糖酸酸合成酶(AS)的缺乏症。在所有新生儿发作性缺乏症(完全酶促缺乏症,在生命的前28天内呈现)中均表示。也有患有高症脑病病史的晚发疾病(部分酶促缺乏症,部分酶促缺乏症,出现)。
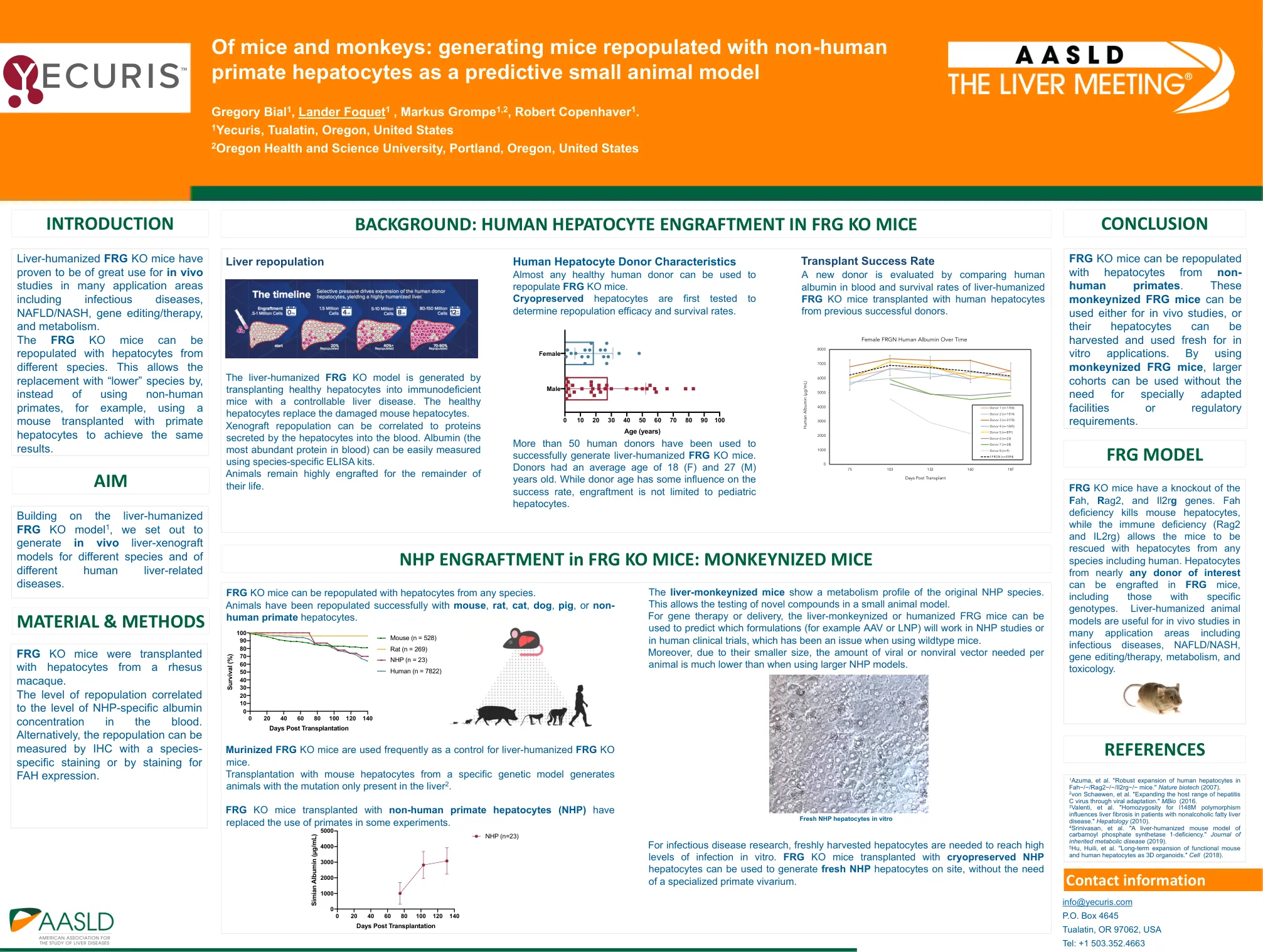
:生成以非人类灵长类动物肝细胞为预测的小动物模型的小鼠
1 Azuma等。“人类肝细胞在fah - / - /rag2 - / - /il2rg - / - 小鼠中的稳健膨胀。”自然生物技术(2007)。2冯·施文(Von Schaewen)等。“通过病毒适应扩大丙型肝炎病毒的宿主范围。”MBIO(2016。 3 Valenti等。 “ I148M多态性的纯合性影响非酒精性脂肪肝病患者的肝纤维化。” Hepatology(2010)。 4 Srinivasan等。 “肝磷酸合成酶1-缺乏的肝脏小鼠模型。” 遗传代谢疾病杂志(2019年)。 5 Hu,Huili等。 “功能小鼠和人肝细胞作为3D器官的长期扩张。” Cell(2018)。MBIO(2016。3 Valenti等。 “ I148M多态性的纯合性影响非酒精性脂肪肝病患者的肝纤维化。” Hepatology(2010)。 4 Srinivasan等。 “肝磷酸合成酶1-缺乏的肝脏小鼠模型。” 遗传代谢疾病杂志(2019年)。 5 Hu,Huili等。 “功能小鼠和人肝细胞作为3D器官的长期扩张。” Cell(2018)。3 Valenti等。“ I148M多态性的纯合性影响非酒精性脂肪肝病患者的肝纤维化。”Hepatology(2010)。 4 Srinivasan等。 “肝磷酸合成酶1-缺乏的肝脏小鼠模型。” 遗传代谢疾病杂志(2019年)。 5 Hu,Huili等。 “功能小鼠和人肝细胞作为3D器官的长期扩张。” Cell(2018)。Hepatology(2010)。4 Srinivasan等。“肝磷酸合成酶1-缺乏的肝脏小鼠模型。”遗传代谢疾病杂志(2019年)。5 Hu,Huili等。 “功能小鼠和人肝细胞作为3D器官的长期扩张。” Cell(2018)。5 Hu,Huili等。“功能小鼠和人肝细胞作为3D器官的长期扩张。”Cell(2018)。Cell(2018)。

使用液滴微流体技术进行可扩展且自动化的基于 CRISPR 的菌株工程
摘要 我们提出了一种基于液滴的微流体系统,该系统可在芯片上实现基于 CRISPR 的基因编辑和高通量筛选。微流体装置包含一个 10 × 10 元件阵列,每个元件包含用于两个电场驱动操作的电极组:用于合并液滴以混合试剂的电润湿和用于转化的电穿孔。该装置可以并行执行多达 100 个基因改造反应,为生成遗传途径组合优化和可预测生物工程所需的大量工程菌株提供了一个可扩展的平台。我们通过基于 CRISPR 的两个测试案例的工程改造展示了该系统的能力:(1)破坏大肠杆菌中酶半乳糖激酶(galK)的功能;(2)靶向改造谷氨酰胺合成酶基因(glnA)和蓝色色素合成酶基因(bpsA),以提高大肠杆菌中的靛蓝素产量。
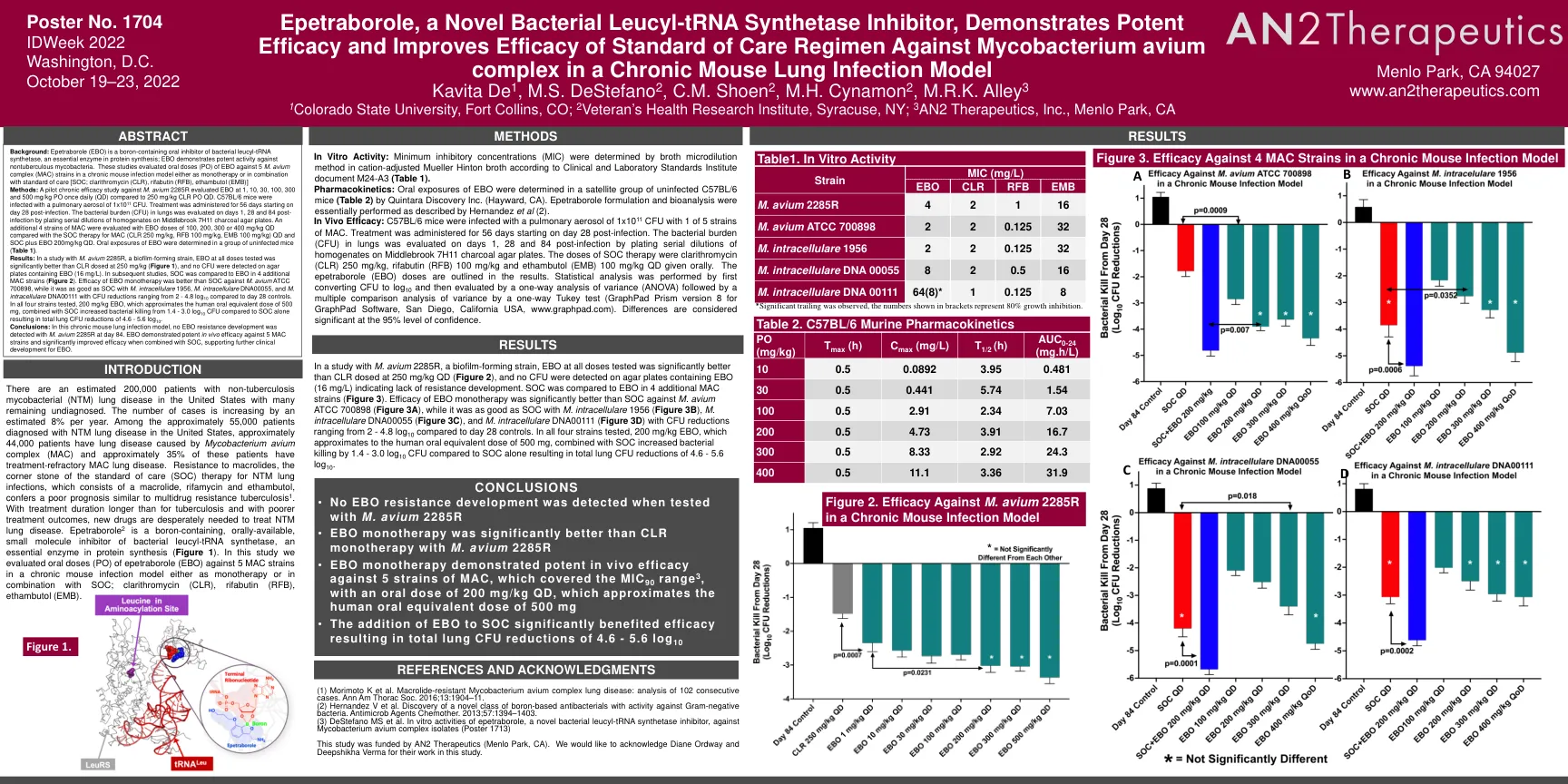
Epetraborole是一种新型的细菌亮氨基-TRNA合成酶抑制剂,它表现出有效的功效并提高了护理标准方案的功效
背景:Epetraborole(EBO)是含硼的口服叶木基-TRNA合成酶的口服抑制剂,这是蛋白质合成中必不可少的酶; EBO表现出对非结核分枝杆菌的有效活性。这些研究评估了EBO的口服剂量(PO)针对慢性小鼠感染模型中的5 M. Avium复合物(MAC)菌株作为单一疗法或与标准护理[SOC;克拉霉素(CLR),利法布丁(RFB),ethambutol(emb)]方法:针对Avium 2285R M. 2285r评估EBO的试验性慢性疗效研究,每天1、10、30、100、300和500 mg/kg PO每天(QD)(QD),而不是250 mg/kg/kg Clr PO QD。C57BL/6小鼠用1x10 11 CFU的肺气溶胶感染。从感染后第28天开始进行56天的治疗。在感染后第1、28和84天评估肺中的细菌负担(CFU),通过在Middlebrook 7H11木炭琼脂板上镀匀性稀释液。与MAC的SOC治疗(CLR 250 mg/kg,RFB 100 mg/kg,100 mg/kg),EBO剂量为100、200、300或400 mg/kg QD评估了4株Mac菌株。在一组未感染的小鼠中确定了EBO的口服暴露(表1)。 结果:在对Avium 2285R的一项研究中,生物膜形成菌株,EBO在所有剂量上测试的EBO明显好于以250 mg/kg剂量的CLR(图1),并且在含有EBO的琼脂平板上检测到NO NO CFU(16 mg/L)。 在随后的研究中,将SOC与其他4种MAC菌株中的EBO进行了比较(图2)。 结论:在这种慢性小鼠肺部感染模型中,在第84天未检测到Avium 2285R的EBO耐药性发展。在一组未感染的小鼠中确定了EBO的口服暴露(表1)。结果:在对Avium 2285R的一项研究中,生物膜形成菌株,EBO在所有剂量上测试的EBO明显好于以250 mg/kg剂量的CLR(图1),并且在含有EBO的琼脂平板上检测到NO NO CFU(16 mg/L)。在随后的研究中,将SOC与其他4种MAC菌株中的EBO进行了比较(图2)。结论:在这种慢性小鼠肺部感染模型中,在第84天未检测到Avium 2285R的EBO耐药性发展。EBO单一疗法的功效比SOC比对Avium ATCC 700898更好,而与M. Intacellulare 1956,M。el. ellacelulare DNA00055和M. el. ellacululare DNA00111相比,与2-4.8 log 10相比,它与M. Intarululare DNA00055和M. M. soc一样好。在测试的所有四种菌株中,200 mg/kg EBO近似于500 mg的人口腔等效剂量,与单独使用SOC相比,SOC的细菌杀死从1.4-3.0 log 10 CFU增加,从而导致总肺CFU降低总量为4.6-5.6 log 10。eBO与5种MAC菌株具有有效的体内功效,并在与SOC结合使用时会显着提高功效,从而支持EBO的进一步临床发育。

CRISPR/Cas9 靶向抑制菊苣中的木香烃内酯合成酶,导致木香烃内酯及其结合物在主根中积累
菊苣主根积累倍半萜内酯乳酸素、乳苦素和 8-脱氧乳酸素,主要以草酸形式存在。菊苣倍半萜内酯的生物合成途径仅部分阐明;将法呢基焦磷酸转化为木香烃内酯的酶已被描述。木香烃内酯转化为三环结构愈创木香烃内酯的下一个生物合成步骤,迄今为止在菊苣中尚未阐明。在这项研究中,在菊苣中发现了三种假定的木香烃内酯合酶基因,分别名为 CiKLS1、CiKLS2 和 CiKLS3。使用酵母微粒体测定法在体外证明了它们将木香烃内酯转化为木香烃内酯的活性。接下来,将 CRISPR/Cas9 试剂引入菊苣原生质体,以灭活多个菊苣 KLS 基因,并成功再生了几个菊苣品系。通过 CRISPR/Cas9 方法灭活菊苣中的 kauniolide 合酶基因,导致菊苣叶和主根中倍半萜内酯的生物合成中断。在菊苣主根中观察到木香烃内酯及其结合物的积累量很高,即 1.5 mg/g FW,但在叶子中没有。这些结果证实,尽管程度不同,但所有这三个基因都有助于 STL 的积累。这些观察结果表明,菊苣基因组上串联的三个基因编码 kauniolide 合酶,可启动菊苣中木香烃内酯向倍半萜内酯的转化。

对硅藻三角褐指藻中的长链酰基辅酶 A 合成酶家族进行多重 CRISPR/Cas9 编辑,发现线粒体 ptACSL3 参与了储存脂质的合成
Xiahui Hao、Wenchao Chen、Alberto Amato、Juliette Jouhet、Eric Maréchal 等人。硅藻 Phaeodactylum tricornutum 中长链酰基辅酶 A 合成酶家族的多重 CRISPR/Cas9 编辑表明,线粒体 ptACSL3 参与了储存脂质的合成。New Phytologist,2022 年,233 (4),第 1797-1812 页。�10.1111/nph.17911�。�hal-03479559�

植物抗癌药物的代谢工程
测序和转录组学的进步使得通过共表达分析可以发现酶,其中候选基因通过组织表达模式与已知途径酶的相似性来识别 — — 最近在 C.roseus 和 Podophyllumpeltatum 中的发现证明了这一点 [ 4 , 5 ]。自组织映射等机器学习方法进一步优化了候选基因 [ 6 ]。这些方法,加上对植物体内生物合成定位的更深入理解,以及单细胞代谢组学等技术的发展,进一步改善了候选基因的选择,加速了酶的发现 [ 7 ]。借助基于 OMIC 的工具(如 plantiSMASH)识别物理基因簇有助于阐明缺失的生物合成酶,如那可丁和长春花碱途径中的酶 [ 8–10 ]。然而,这种方法是有限的,因为许多植物生物合成途径几乎没有或没有基因聚集,如喜树碱生物合成途径[11]。基于同源性的克隆可以加速发现与已知生物合成酶具有直系同源功能的基因,例如在 Tabernanthe iboga 的 ibogaine 生物合成途径中鉴定出 C. roseus 脱羧酶直系同源物[12]。然而,途径的复杂性往往需要采用组合方法,例如 Gelsemiumsempervirens 氧化吲哚途径的发现[13]。

16S rRNA测序和靶向代谢组学分析揭示了根际微生物的多样性和类黄酮的动力学,在Baicalaria Baicalaria georgi
黄金中黄酮的生物合成途径已被广泛阐明,主要通过根特异性的黄酮途径(Fang等人。2022)。gente异黄酮合成途径起源于肉桂酸(图1),在SBPAL的作用下从氨基酸苯丙氨酸合成为生物合成前体。肉桂酸随后通过cinnamoyl coa连接酶转化为肉桂酸COA。pine chalcone合成酶催化肉桂酸COA产生pinocembrin chalcone,该核蛋白结构蛋白通过chalcone异构酶进行异构化,以产生pinocembrin。然后,类黄酮合成酶将pinocembrin转换为chrysin,该酸蛋白被6-羟化酶进一步羟基羟基羟基酶(Liu et al。2021)。黄氨基蛋白是由Baicalin-7-O-葡萄糖糖基转移酶葡萄糖醛酸糖苷至Baicalin,而Chrysin则被F8H转化为Norwogonin。NORWOGONIN通过O-甲基转移酶(OMT)在位置8的位置进行O-甲基化,以产生Wogonin,最终通过Baicalin-7-O-o-葡萄糖糖基转移酶将其葡萄糖醛酸化为Wogonoside(Pei等人。 2023)。NORWOGONIN通过O-甲基转移酶(OMT)在位置8的位置进行O-甲基化,以产生Wogonin,最终通过Baicalin-7-O-o-葡萄糖糖基转移酶将其葡萄糖醛酸化为Wogonoside(Pei等人。2023)。