XiaoMi-AI文件搜索系统
World File Search System合成酶

16S rRNA测序和靶向代谢组学分析揭示了根际微生物的多样性和类黄酮的动力学,在Baicalaria Baicalaria georgi
黄金中黄酮的生物合成途径已被广泛阐明,主要通过根特异性的黄酮途径(Fang等人。2022)。gente异黄酮合成途径起源于肉桂酸(图1),在SBPAL的作用下从氨基酸苯丙氨酸合成为生物合成前体。肉桂酸随后通过cinnamoyl coa连接酶转化为肉桂酸COA。pine chalcone合成酶催化肉桂酸COA产生pinocembrin chalcone,该核蛋白结构蛋白通过chalcone异构酶进行异构化,以产生pinocembrin。然后,类黄酮合成酶将pinocembrin转换为chrysin,该酸蛋白被6-羟化酶进一步羟基羟基羟基酶(Liu et al。2021)。黄氨基蛋白是由Baicalin-7-O-葡萄糖糖基转移酶葡萄糖醛酸糖苷至Baicalin,而Chrysin则被F8H转化为Norwogonin。NORWOGONIN通过O-甲基转移酶(OMT)在位置8的位置进行O-甲基化,以产生Wogonin,最终通过Baicalin-7-O-o-葡萄糖糖基转移酶将其葡萄糖醛酸化为Wogonoside(Pei等人。 2023)。NORWOGONIN通过O-甲基转移酶(OMT)在位置8的位置进行O-甲基化,以产生Wogonin,最终通过Baicalin-7-O-o-葡萄糖糖基转移酶将其葡萄糖醛酸化为Wogonoside(Pei等人。2023)。

具有光子计数检测器CT的钙化冠状动脉中的虚拟钙去除 - 首先体验体验
在1993年,日本研究人员首先报道了患有临床和生物化学疾病的成年患者,类似于尿素周期酶Argininoscinate合成酶1的缺陷引起的经典柑橘类血症1型,但在相应的ASS1基因中缺乏遗传变异。1同一位作者报道了这种情况,称为Citrullinemia类型2或CTLN2,其特征是肝氨基辛酸核酸酯合成酶1(ASS1)的降低,具有正常的动力学特性和热稳定性,伴随着接近正常水平的Ass1 mRNA肝脏中的肝脏中的肝脏1 mRNA,肝脏中的正常水平,正常的翻译活动,没有正常的翻译结构,没有毛的结构效果。1最后,Kobayashi等。将CTLN2的原则确定为不是源自ASS1基因座的,并成功地克隆了因子基因SLC25A13,为2,它们指定为“ Citrin一词”。”基于这种历史的观点,现在被称为由SLC25A13突变引起的常染色体隐性疾病β-氧化,三羧酸(TCA)周期和尿素周期。The disease is characterized by age- dependent, variable clinical manifestations: neonatal intrahepatic cholestasis caused by citrin deficiency (NICCD; OMIM 605814), failure to thrive and dyslipide- mia caused by citrin deficiency (FTTDCD), and adult- onset type II citrullinemia (CTLN2; OMIM 603471).3 - 5

海报摘要
1个生命科学学院,Jawaharlal Nehru大学,印度新德里,美国亚特兰大2号埃默里大学[P3]基于植物化学的抗淀粉样蛋白银纳米颗粒Om Prakash Mahato Mahato,Kailash Pd。prajapati,Bibin g anand,Shikha Mittal,M。Ansari,Karunakar Kar Life Sciences,Jawaharlal Nehru大学,印度新德里,印度新德里[P4] C. bictyaltransement and sudha kyhha,Sudha,Sudha,Sudhha,Susran sharha,Sudha sharmath sharha,萨穆德拉拉·古林纳特(Samudrala Gourinath)科学,印度新德里的贾瓦哈拉尔·纳赫鲁大学[P6]氨基酰基-TRNA合成酶:药物发现的基本目标,阿米特·沙尔马分子医学 - 结构性寄生虫学小组,国际基因工程和生物技术中心,基因工程和生物技术中心D-Crystallin在印度新德里的Jawaharlal Nehru大学Rajesh Mishra生物技术学院的酸性和生理pH Indu [P8]疟疾和生化描绘疟疾苯丙氨酸苯胺-TRNA合成酶(FRS)。Nachiappan Mutharasappan,Yogavel Manickam,Jyoti Chhibber-Goel,Amit Sharma分子医学 - 结构性寄生虫学小组,国际基因工程与生物技术中心,新德里,印度,

自适应景观平坦化允许设计两种酶:底物结合和催化能力
设计酶具有基础和技术意义。实验定向进化仍然有很大的局限性,计算方法是一条补充途径。设计的酶应满足多个标准:稳定性、底物结合、过渡态结合。这种多目标设计在计算上具有挑战性。最近的两项研究使用自适应重要性抽样蒙特卡罗重新设计蛋白质以进行配体结合。通过首先平坦化载脂蛋白的能量景观,他们获得了结合状态的正设计和非结合状态的负设计。我们现在已将该方法扩展到设计一种酶以进行特定的过渡态结合,即其催化能力。我们考虑了甲硫氨酰-tRNA 合成酶 (MetRS),它将甲硫氨酸 (Met) 附着到其同源 tRNA 上,从而建立密码子身份。此前,MetRS 和其他合成酶已通过实验定向进化重新设计,以接受非规范氨基酸作为底物,从而导致遗传密码扩展。在这里,我们通过计算重新设计了 MetRS,使其能够结合多种配体:Met 类似物叠氮亮氨酸、甲硫氨酰腺苷酸 (MetAMP) 以及形成 MetAMP 生成过渡态的活化配体。通过设计计算恢复了已知具有叠氮亮氨酸活性的酶突变体,并对预测结合 MetAMP 的 17 种突变体进行了实验表征,发现它们均具有活性。预测具有低活化自由能的突变体在 MetAMP 生成中被发现具有活性,并且预测的反应速率与实验值非常吻合。我们建议本方法应成为计算酶设计的范例。

植物生物技术。24.0205a(2024)
在特定的植物家族中发现了抽象的结构上多样化的苄基喹啉生物碱(偏见),其中一些是由于其强大的生物学活性而有效产生的。corydalis植物(例如,Corydalis Yanhusuo)的豆豆科也会产生各种偏见;因此,它们已用于中医。由于使用微生物的代谢工程和合成生物学是有效生产有用代谢物的有前途的技术,因此阐明每个BIA的生物合成途径是必不可少的。尽管最近已经分离出了几种参与corydalis偏差的生物合成的酶基因,但许多酶基因仍然未知,例如蛋白酶碱Calkaloid c-甲基转移酶参与Corydaline的生物合成,这是Corydaline的生物合成,这是Corydalis植物中发现的主要偏见之一。在这项研究中,我们进行了转录组分析,并结合了Corydalis solida不同组织的代谢物分析。基于多种偏差的高积累,包括原生质,同种丙蛋白酶和corydaline,编码推定的生物合成酶的基因,包括细胞色素P450,甲基转移酶和氧化酶蛋白,在块茎中高度表达。在块茎中高度表达了两个OMT基因CSOMT1和CSOMT2,并使用粗酶制剂进行了进一步的表征,表明CSOMT1表现出7- O-甲基化活性,针对网状,而CSOMT2 2催化9- O-甲基化的Sculerine scoulerine,然后由2- o甲基化。我们的发现提供了有价值的信息,用于隔离Corydalis物种中新型生物合成酶基因。

回顾辅酶A及其衍生物的代谢在疾病中起着至关重要的作用
辅酶A(COA)充当细胞内酰基的关键载体,在调节酰基转移反应并参与细胞代谢过程中起着基本作用。作为主要底物和辅助因子从事各种代谢反应,COA及其衍生物对各种生理过程产生了中心影响,主要是调节脂质和酮代谢以及蛋白质修饰。本文对COA的分子机制进行了全面综述,该机制会影响癌症的发作和进展,心血管疾病(CVD),神经退行性疾病和其他疾病。主要焦点包括以下内容。(1)在癌症中,诸如乙酰-COA合成酶2,ATP柠檬酸裂解酶和乙酰辅酶A羧化酶等酶通过调节乙酰-COA水平调节脂质合成和能量代谢。(2)在CVD中,诸如稳态 - coA脱发酶-1、3-羟基-3-羟基-3-甲基戊二核-COA(HMGC)合成酶2和HMGC还原酶的影响以及这些疾病的形成和进步是由Coa Metbolism跨多orgbolism跨越了这些疾病的形成和进步。(3)在神经退行性疾病中,COA在维持大脑中胆固醇稳态及其对此类疾病发展的影响方面的意义得到了详尽的讨论。涉及COA及其衍生物的代谢过程涵盖了细胞内的所有生理方面,在各种疾病的发作和进展中起关键作用。阐明COA在这些疾病中的作用会产生重要的见解,这些见解可以作为疾病诊断,治疗和药物开发的有价值的参考和指导。
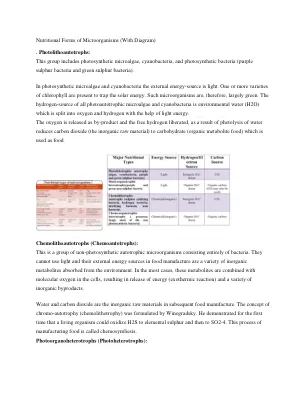
微生物的营养形式(附图)
生长曲线 生长曲线是绘制细胞数量与时间因子的关系图。将已知浓度的细菌接种到合适的培养基(分批培养)中,即可获得典型的生长曲线。细菌通过二分裂生长。每隔一小时定期计数细菌细胞。绘制细菌数量与时间的关系图。典型的细菌生长曲线显示四个不同的阶段,即滞后期、对数期、稳定期和衰退期。滞后期代表细胞数量增加的初始阶段。在此阶段,细胞代谢活跃,能够修复细胞损伤并合成酶。

蛋氨酸代谢酶的基因检测,包括MTHFR
甲基肾上腺酸还原酶(MTHFR),蛋氨酸合酶(MTR),蛋氨酸合酶还原酶(MTRR),钴胺素还原酶(MMADHC)(MMADHC)和胱硫醇β-合成酶(CBS)是提供指导的基因在将氨基酸同型半胱氨酸(HCY)转换为蛋氨酸方面发挥作用。当存在基因的异常拷贝时,它们可能导致酶功能降低,导致同型半胱氨酸水平升高。血液中异常高水平的HCY与几种慢性疾病有关,例如注意力缺陷/多动症(ADHD),心血管疾病,癫痫,头痛,胃肠道症状和状况,精神疾病,精神疾病,骨质疏松症和骨质疾病。

动质体原生生物中不同的细胞色素c成熟系统
摘要 在真核生物中,血红素通过两个硫醚键附着到线粒体细胞色素 c 和 c 1 上,由多亚基细胞色素 c 成熟系统 I 或全细胞色素 c 合成酶 (HCCS) 催化。前者是从线粒体的 α 变形菌祖先遗传而来;后者是一种真核创新,其原核祖先并不明显。HCCS 是真核生物中从头蛋白质创新的少数几个例子之一,但对 HCCS 的结构功能了解有限。独特的是,眼虫原生生物(包括医学上相关的动基体锥虫和利什曼原虫寄生虫)通过单个硫醚键将血红素附着到线粒体 c 型细胞色素上。但该机制尚不清楚,因为缺乏编码与其他分类群中参与细胞色素 c 成熟的蛋白质具有可检测相似性的蛋白质的基因。在这里,通过生物信息学搜索所有含血红素蛋白的动质体中保守的蛋白质,鉴定出动质体细胞色素 c 合成酶 (KCCS),我们发现它是必需的和线粒体的,能催化血红素附着到锥虫细胞色素 c 上。KCCS 与其他蛋白质没有序列同一性,除了四个短基序内的轻微相似性表明与 HCCS 相关。因此,KCCS 为研究真核细胞色素 c 成熟提供了一种新的资源,可能具有更广泛的相关性,因为人类 HCCS 的突变会导致疾病。此外,与许多其他真核生物相比,眼虫的许多线粒体生物化学例子都不同;因此,KCCS 的鉴定为进化分化的原生生物群体中极端、不寻常的线粒体生物化学提供了另一个典范。

复活的cas12a祖先扩展了对核酸编辑和检测底物的目标和识别的访问
据众所周知,RECHB是唯一描述的具有这种扩展活性的核酸酶。 div>很有可能在自然界中具有这些特征,但是在天然酶的空间中,可能会很艰巨,昂贵且需要很长时间。 div>同样,基于自动学习的计算方法仍在开始,尚无法设计具有复杂和受控功能的酶,例如大型构象变化。 div>开发了深度学习方法(OpenCrispri-1),尽管有希望,但尚未证明具有新功能设计蛋白质的能力。 div>这些限制突出了ASR生成具有多种和改进特性的复杂合成酶的能力,并开放了与深度学习和语言方法结合的新方法。 div>