XiaoMi-AI文件搜索系统
World File Search System密码子

卫生技术简报2023年5月
NSCLC是英国最常见的肺癌形式,其亚型可以定义为腺癌,鳞状细胞癌和大细胞癌。10转移性NSCLC(第4阶段)是癌症扩散到人体其他部位的时候,例如骨骼,肺,脑,肝或肾上腺。11局部晚期NSCLC(第3A期)是癌症,已扩散到肺周围的组织中。12 KRAS G12C突变是最常见的,发生在英国12%的非小细胞肺癌(NSCLC)肿瘤中。13 A KRAS G12C突变是KRAS基因密码子12的甘氨酸残基的突变。这种突变导致RAS信号传导的激活,这可能导致自发的肿瘤发育和肿瘤微环境的产生,从而使肿瘤的增殖和维持。14肺癌最常见的症状包括咳嗽,呼吸困难,咳嗽,血液疼痛,胸部或肩膀疼痛,胸部反复感染,食欲不振,体重减轻和疲劳。15一些研究表明,吸烟状况与不同的KRAS突变和密码子变体的关联。与非KRAS G12C突变或野生型KRAS相比,来自手术切除的肺腺癌的KRAS G12C突变患者的2年总生存率明显较差。16

噬菌体WO基因wmk的杀伤表型
摘要沃尔巴克氏体是动物体内分布最广泛的细菌内共生体。在节肢动物中,这些母系传播的细菌可以自私地劫持宿主的生殖过程,以提高传播它们的雌性的相对适应度。一种称为雄性杀灭或选择性杀死受感染雄性的生殖寄生形式,通过转基因表达原噬菌体 WO 介导的杀灭 ( wmk ) 基因在一定程度上重现。在这里,我们利用转基因表达表征了果蝇中 wmk 诱导的雄性杀灭的基因型-表型景观。虽然系统发育上遥远的 wmk 同源物不会引起性别比例偏差,但密切相关的同源物表现出复杂的表型,包括不死亡、雄性死亡或所有宿主死亡。我们证明替代起始密码子、同义密码子,尤其是 wmk 中的单个同义核苷酸可以消除杀灭。这些发现揭示了转基因 wmk 诱导杀戮的先前未被认识到的特征,并为转录后过程对雄性杀戮变异的影响建立了新的假设。我们得出结论,同义序列变化在具有生死后果的嵌套内共生相互作用中不一定是沉默的。

社论:转移RNA的合成生物学和治疗应用
在所有活细胞中,遗传代码定义了蛋白质编码基因中核酸序列与需要准确生产基因组中所有蛋白质所需的氨基酸序列之间的关系。氨基酰基-TRNA合成酶对转移RNA(TRNA)的氨基酰化是将氨基酸与TRNA守流量将氨基酸物理联系起来的关键步骤,从而决定了密码子与氨基酸的分配。由于它们在蛋白质合成中的核心作用,设计和合成的TRNA作为正交翻译系统的重要组成部分,旨在将非典型甚至不自然的氨基酸掺入细胞和无细胞系统中的蛋白质中。此外,最近的努力使用了正常的野生型或工程性TRNA来纠正引起人类疾病的遗传缺陷。由于11%的遗传遗传疾病是由过早的停止密码子引起的,因此废话抑制剂TRNA对tRNA疗法的应用越来越兴趣。我们认识到,TRNA的合成生物学和治疗应用都将依赖于胡说八道,在某些情况下,错过抑制tRNA,生成新型蛋白质或纠正遗传缺陷。因此,遗传学领域的研究主题具有遗传代码扩展和探索TRNA在合成生物学和医学应用中的应用中的作用的研究。

CodeBreak 200:研究局限性和未来方向
近年来,肺癌研究的突破性进展为直接针对 KRAS 突变的创新治疗铺平了道路。长期以来,KRAS 突变一直被认为无法用药,因为它们对底物 [鸟苷三磷酸 (GTP)] 的亲和力高,达到皮摩尔水平,且缺乏已知的调节结合位点。非小细胞肺癌 (NSCLC) 中的 KRAS 突变最常发生在外显子 2 和 3 的热点中,特别是在密码子 12、13 和 61 处,不同突变体的生化特性各不相同 (1)。值得注意的是,KRAS p.G12C 突变的特征是第 12 个密码子上的甘氨酸 (G) 被半胱氨酸 (C) 取代,约占非小细胞肺癌中发现的 KRAS 突变的 40%,或西半球非小细胞肺癌中已知致癌驱动因素的 13-16%,与其他 KRAS 突变相比,其内在 GTP 水解水平接近正常水平,因此能够在鸟苷二磷酸 (GDP) 结合(失活)和 GTP 结合(活性)状态之间循环。除了结构分析方面的进步外,这种基因型特异性的生化特征为产生突变选择性共价抑制剂奠定了基础,这种抑制剂可以不可逆地与 GDP 结合(失活)形式的 KRAS G12C (2) 结合。

在Lynch综合征中识别出的罕见的MLH1重复
结果:在MMR基因中,在DHPLC基因变异筛选中鉴定了三个家族,该家族在MMR基因中具有致病性/可能的致病性种系变体。所有家庭在几代人的几个家庭成员中都有CRC的历史。肿瘤分析表明,与突变基因以及MSI相对应的MMR蛋白IHC染色的丢失。在MLH1中鉴定出a的a family A,一种结构变体,4至13的重复。预计重复将导致氨基酸520的框架和氨基酸539的过早终止密码子。在家庭B中,在MLH1中发现了1个碱基对缺失,从而在氨基酸491中产生移牌和终止密码子。在家庭C中,我们确定了MSH2中的一个剪接位点变体,该变体预计将导致剪接供体部位的损失。结论:我们在19个测序家族中的三个中,在MMR基因中完全确定了三种致病/可能的致病变异。基于洞察力和Clinvar数据库,MLH1变体是外显子4至13的复制和移码变体的新颖。 Clinvar的一个提交者报告了MSH2剪接网站变体。作为一种变体类别,在MMR基因文献中很少报道重复,尤其是涵盖多个外显子的文献。
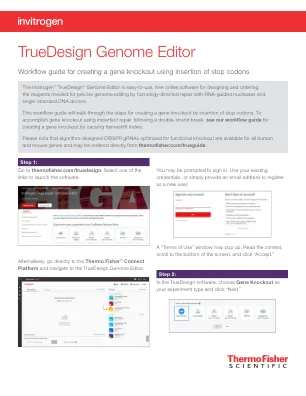
TrueDesign 基因组编辑器
视图将更新以显示“选择 gRNA 选项”。单击适合您需求的 sgRNA 选项 - “预先设计的 sgRNA”(仅限人类和小鼠基因)或“自定义 gRNA 和 TALEN 设计”。如果您对插入终止密码子的区域没有偏好,并且正在人类或小鼠细胞中工作,建议使用“预先设计的 sgRNA”。否则,选择“自定义 gRNA 和 TALEN 设计”。本文档中显示的大多数步骤对于两个工作流程都相似。

ans-Assessment-Questions.pdf
以下是密码子学习的自主神经系统单元的评估问题,您可以在上课前或上课前测试学生的知识。每个问题后都给出了正确的答案和Bloom的分类级别。访问www.codonlearning.com,以了解我们的高结构解剖学和生理学课程,该课程由贾斯汀·沙弗(Justin Shaffer)在科罗拉多州矿业学院设计,该课程使用循证教学实践来改善学生的学习和成果。1。由于交感神经导致以下哪种影响?

利用 TadA 的下一代胞嘧啶碱基编辑器
• 脱氨酶的定向进化 • PAM 变体碱基编辑器 • 定向进化 Cas9 以创建用于 BE 的非 NGG PAM 变体 • 密码子、NLS 和接头优化 • 环状置换体和镶嵌碱基编辑器 • DNA 脱靶评估 • RNA 脱靶评估 • 旁观者编辑最小化 • 引导 RNA 工程 • 离体和体内 BE 递送 • 最小化脱靶活性的工程 BE • HSC、肝细胞和 T 细胞的离体碱基编辑 • ABE 的低温电子显微镜结构 • 小鼠体内碱基编辑 • 非人类灵长类动物体内编辑

最初发表于:Heeb,Laura V;塔斯科帕兰,贝图尔;卡特苏拉斯(Katsoulas),安东尼奥斯(Antonios);贝芬格,米哈尔;克拉维恩,皮埃尔-阿兰;狗头人,塞巴斯蒂安;古普塔,
免疫抑制分子程序性细胞死亡配体 1 (PD-L1) 已被证明在自身免疫、感染和癌症等病理中发挥作用。PD-L1 不仅在癌细胞上表达,而且在未转化宿主细胞上的表达也与癌症进展有关。小鼠系统中 PD-L1 缺陷的产生使我们能够专门研究 PD-L1 在生理过程和疾病中的作用。最通用且最易于使用的位点特异性基因编辑工具之一是 CRISPR/Cas9 系统,它基于 RNA 引导的核酸酶系统。与其前身锌指核酸酶或转录激活因子样效应核酸酶 (TALEN) 类似,CRISPR/Cas9 催化双链 DNA 断裂,这可能导致由于非同源末端连接 (NHEJ) 的随机核苷酸插入或缺失而导致的移码突变。此外,尽管不太常见,但 CRISPR/Cas9 可以在存在合适模板的情况下通过同源定向修复 (HDR) 导致插入确定的序列。在这里,我们描述了使用 CRISPR/Cas9 在小鼠 C57BL/6 背景下敲除 PD-L1 的方案。外显子 3 的靶向结合 HindIII 限制位点的插入会导致过早终止密码子和功能丧失表型。我们描述了靶向策略以及创始者筛选、基因分型和表型。与基于 NHEJ 的策略相比,所提出的方法可产生具有与 NHEJ 相当的效率和时间线的确定终止密码子,生成方便的创始者筛选和基因分型选项,并且可以快速适应其他目标。

酿酒酵母表达系统代谢调控的先进技术和新进展展望
酿酒酵母是广泛使用的生物合成系统之一,用于生产各种生物产品,尤其是生物治疗药物和重组蛋白。由于外来基因的表达和插入总是受到酿酒酵母内源性因素和非生产性程序的阻碍,因此已经开发出各种技术来增强转录的强度和效率并促进基因编辑程序。因此,阻碍异源蛋白质分泌的限制已经得到克服。已经开发出负责转录起始和精确调控表达的高效启动子,这些启动子可以通过合成启动子和双启动子表达系统进行精确调控。适当的密码子优化和协调以适应酿酒酵母的基因组密码子丰度有望进一步提高转录和翻译效率。通过将专门设计的信号肽与上游外源基因融合,可以实现高效、准确的转运,从而促进新合成的蛋白质的分泌。除了广泛应用的启动子工程技术和明确的内质网分泌途径机制外,创新的基因组编辑技术 CRISPR/Cas(成簇的规律间隔的短回文重复序列/CRISPR 相关系统)及其衍生工具可以更精确、更有效地进行基因破坏、定点突变和外源基因插入。本综述重点介绍为精确调控酿酒酵母表达系统的代谢而开发的复杂工程技术和新兴遗传技术。