XiaoMi-AI文件搜索系统
World File Search System肺泡

在遗传性肺肺泡蛋白质病的鼠模型中,有效的造血干细胞基因疗法
1德国汉诺威汉诺威医学院实验血液学研究所; 2德国汉诺威汉诺威医学院功能和应用解剖研究所; 3美国俄亥俄州辛辛那提市辛辛那提儿童医院医疗中心肺部生物学系转换肺科学中心; 4日本Tochigi的Shimotsukeshi Jichi医科大学肺医学系; 5德国汉诺威汉诺威医学院放射学系; 6德国汉诺威汉诺威医学院核医学系; 7美国俄亥俄州辛辛那提市辛辛那提儿童医院医疗中心实验血液学和癌症生物学,癌症与血液疾病研究所(CBDI); 8美国马萨诸塞州波士顿哈佛医学院波士顿儿童医院血液学/肿瘤学系; 9汉诺威医学院,汉诺威,德国汉诺威医学院的儿科,过敏症和新生儿学系,俄亥俄州辛辛那提儿童医院医疗中心肺部医学部门10

文章训练的肺泡巨噬细胞的免疫力通过KLF4-MERTK介导的肿瘤细胞增强了损伤
最近的研究表明,通过反复的有害刺激对先天免疫细胞(例如居住的巨噬细胞)进行培训可以增强宿主防御反应。然而,尚不清楚训练有素的组织居民巨噬细胞的免疫力是否还可以增强损伤的解决方案,以抵消增强的炎症反应。在这里,我们研究了肺部肺泡巨噬细胞(AMS),这些巨噬细胞(AMS)先于细菌内毒素或铜绿假单胞菌,并观察到这些受过训练的AMS对病原体诱导的细胞死亡的韧性更大。转录组分析和功能分析表明,受过训练的AM能力更大,用于细胞碎片和损伤分辨率。单细胞高维质量细胞仪分析和谱系跟踪表明,训练会诱导Mertk Hi Marco HI CD163 + F4/ 80低肺居民AM子集的扩展,并具有预处理表型。重编程的AMS上调了由转录因子KLF4介导的肿瘤受体MERTK的表达。这些受过训练的AMS受限制的炎症性肺损伤的收养转移在暴露于致命的铜绿假单胞菌的受体小鼠中。因此,我们的研究确定了一部分受过组织训练的巨噬细胞,这些巨噬细胞在反复发生病原体挑战后防止过度炎症并恢复组织稳态。

mRNA 疗法——空间传递
5 1. 1 型肺泡细胞 2. 2 型肺泡细胞 3. B 细胞 4. T 细胞和 NK 细胞 5. 肺泡巨噬细胞 6. 成纤维细胞(1 类) 7. 成纤维细胞(2 类) 8. 内皮细胞 9. 间质巨噬细胞 10. 单核细胞 11. 中性粒细胞 12. 周细胞(1 类) 13. 周细胞(2 类)

利用下一代肺泡 2 型类器官平台模拟肺腺癌的多种遗传亚型
肺癌是全球癌症相关死亡的主要原因。肺腺癌 (LUAD) 是最常见的组织学亚型,占所有病例的 40%。虽然现有的基因工程小鼠模型 (GEMM) 重现了人类 LUAD 的组织学进展和转录进化,但它们耗时且技术要求高。相比之下,细胞系移植模型快速灵活,但这些模型无法捕捉疾病进展的全部范围。类器官技术提供了一种创建下一代癌症模型的方法,该模型整合了自体系统和基于移植的系统的最有利特征。然而,目前缺乏强大而可靠的 LUAD 类器官平台。在这里,我们描述了在类器官培养中持续扩增小鼠肺泡 2 型 (AT2) 细胞(LUAD 的主要起源细胞)的优化条件。这些类器官表现出 AT2 细胞的典型特征,包括标记基因表达、层状体的存在以及分化为 AT1 谱系的能力。我们利用该系统开发了灵活且多功能的免疫功能正常的类器官模型,用于 KRAS 、 BRAF 和 ALK 突变型 LUAD。值得注意的是,类器官肿瘤表现出广泛的负担和完全渗透性,并且在组织病理学上与原发肿瘤没有区别。总之,该类器官平台是一个功能强大、用途广泛的新型 LUAD 研究模型系统。

层次药物释放设计AU @PDA-PEG-MTX NP,用于通过合并光热化学疗法靶向乳腺癌的靶向递送
整齐地排列,并且可以接受管状和间质互化结构。au @pda-peg-mtx nps组中glomeruli的体积和大小不一致。肾小球中的细胞比正常人增加,细胞外基质的增加比正常情况大,并且肾小管上皮细胞的排列不规则。肾小管的结构尚不清楚。NIR+AU @PDA-PEG-MTX NPS组与对照组相似。在对照组和两个实验组中,肺组织结构相对清晰,整个肺泡结构相对完整,肺泡壁的厚度相对正常,支气管狭窄的程度相对轻。肺泡上皮细胞,嗜酸性粒细胞和淋巴细胞很少浸润

使用新型的toclock干涉法
压力,细胞连接的破坏以及细胞骨架结构的破坏都可以参与此过程。由活性氧介导的凋亡(ROS)可以通过PI3K/AKT/NFKB/MMP-9癌症中的EMT作用[37]。在COPD中,ROS可以促进上皮表型转化,从而导致上皮细胞的异常增殖和分化,从而导致上皮下胶原蛋白沉积[38]。当前的研究观察到CS可以导致肺泡和气道上皮的EMT。cs可以通过Wnt/β-Catenin信号通路促进肺泡上皮细胞中的EMT,从而导致肺泡修复能力受损[39]。COPD始于小气道功能障碍,因此我们的研究着重于COPD中小气道和气道上皮的变化。
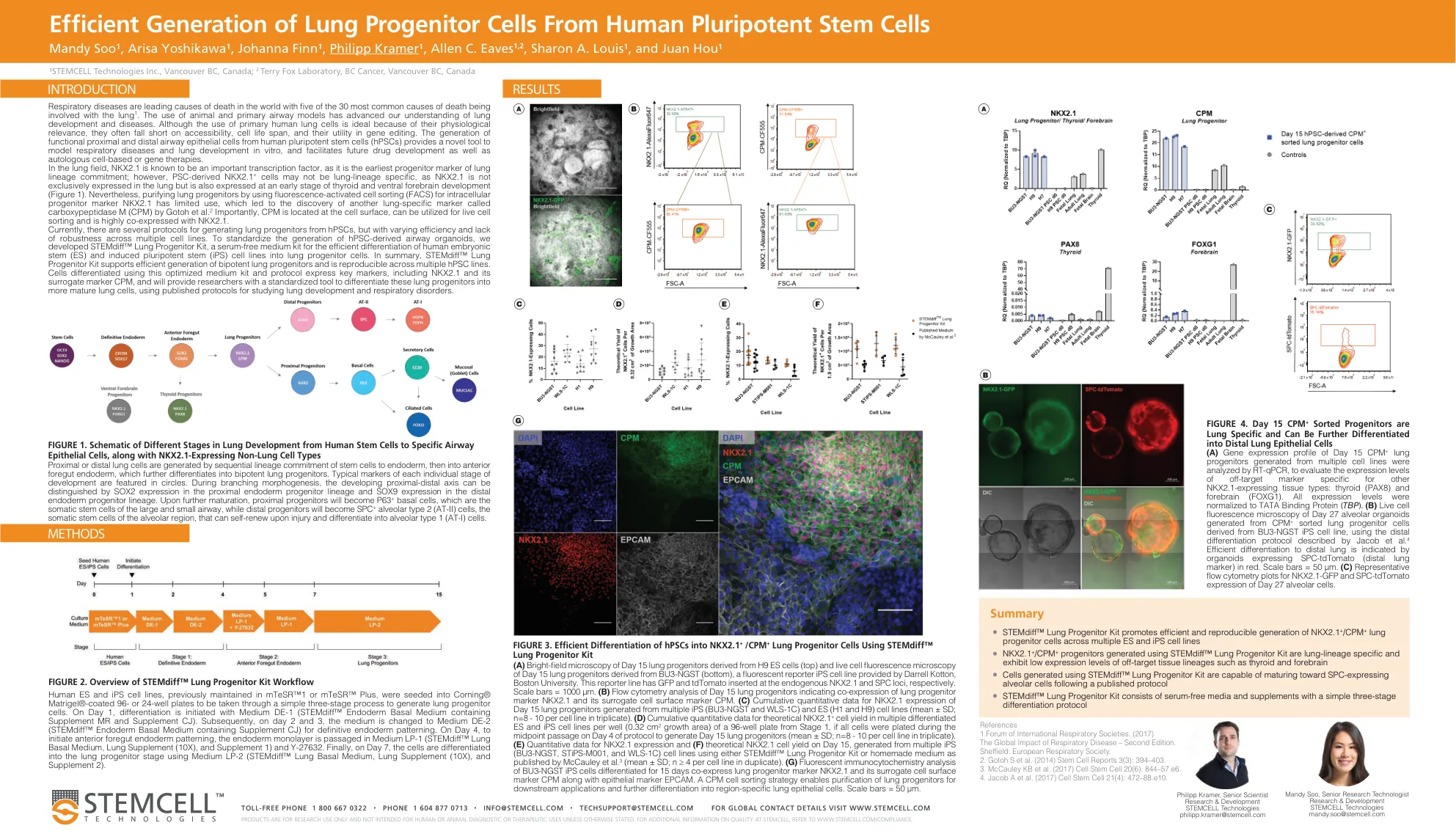
SP00248-从人类多能干细胞高效生成肺祖细胞
近端或远端肺细胞是由干细胞按顺序谱系分化到内胚层,然后进入前肠内胚层,进一步分化为双能肺祖细胞而产生的。每个发育阶段的典型标记以圆圈表示。在分支形态形成过程中,可以通过近端内胚层祖细胞谱系中的 SOX2 表达和远端内胚层祖细胞谱系中的 SOX9 表达来区分发育中的近端-远端轴。进一步成熟后,近端祖细胞将变成 P63 + 基底细胞,即大气道和小气道的体细胞干细胞,而远端祖细胞将变成 SPC + 肺泡 2 型 (AT-II) 细胞,即肺泡区域的体细胞干细胞,它们在受伤后可以自我更新并分化为肺泡 1 型 (AT-I) 细胞。


感染前的先天免疫减轻了IPSC衍生的肺泡上皮2细胞中SARS-COV-2感染和病毒载荷
摘要:2019年冠状病毒病(COVID-19)疾病的易感性和疾病严重程度(SOI)的一大部分异质性仍然知之甚少。最近的证据表明,SARS-COV-2感染相关的损伤对肺部肺部肺泡2型细胞(AT2S)的损伤可能直接导致COVID-19患者的疾病严重程度和预后不良。我们在诱导的多能干细胞(IPSC)衍生的10个不同个体中SARS-COV-2感染的体外模型表现出感染敏感性和感染后细胞病毒载荷的个体变异性。为了了解调节SARS-COV-2感染和细胞病毒载量的AT2'的能力的潜在机制,对模拟和SARS-COV-2感染挑战的AT2S进行了全基因组差异基因表达分析。The 1393 genes, which were significantly (one-way ANOVA FDR-corrected p ≤ 0.05; FC abs ≥ 2.0) differentially expressed (DE), suggest significant upregulation of viral infection-related cellular innate immune response pathways ( p -value ≤ 0.05; activation z -score ≥ 3.5), and significant downregulation of the cholesterol- and xenobiotic-related代谢途径(P值≤0.05;激活Z -SCORE≤ -3.5)。Whilst the effect of post-SARS-CoV-2 infection response on the infection susceptibility and postinfection viral load in AT2s is not clear, interestingly, pre-infection (mock-challenged) expression of 238 DE genes showed a high correlation with the postinfection SARS-CoV-2 viral load (FDR-corrected p -value ≤ 0.05 and r 2 - absolute ≥ 0.57).表达与病毒负荷负相关的85个基因在病毒识别和细胞因子介导的先天免疫GO生物学过程中表现出显着富集(P值范围:4.65×10-10-10-10至2.24×10-6)。表达与病毒载量正相关的153个基因在胆固醇稳态,细胞外基质和MAPK/ERK途径相关的GO生物学过程中表现出显着富集(P -Value范围:5.06×10 - 5至6.53×10-4)。总的来说,我们的结果强烈表明AT2的感染前的免疫和代谢状态会影响其对SARS-COV-2感染和病毒负荷的敏感性。

EGFR驱动的肺癌细胞的细胞起源确定对治疗的敏感性EGFR驱动的肺癌细胞的细胞起源确定对治疗的敏感性
用酪氨酸激酶抑制剂(TKIS)靶向表皮生长因子受体(EGFR)是肺腺癌的主要精度医学治疗选择之一。由于对第一代和第二代TKI的耐药性的共同发展,包括Osimertinib和Rociletinib在内的第三代抑制剂。在这里描述了通过3D器官培养物开发不同表观遗传态肿瘤的EGFR驱动的肺癌模型。发现,肺上皮细胞中EGFR T790M/L858R突变的激活可以驱动具有肺泡或支气管元素的肺癌,这可能源自肺泡2(AT2)细胞或支气管肺泡干细胞,而不是基质细胞,而不是基底细胞或杆基细胞或杆细胞。也证明,这些克隆能够通过在小鼠中的原位传播,至关重要的是,它们具有独特的药物脆弱性,可以保留其表观遗传差异。这项工作是探索如何使用表观遗传学来对患者进行精确医学决策进行分层的蓝图。