XiaoMi-AI文件搜索系统
World File Search SystemMDM2

Milademetan 是治疗默克尔细胞癌的高效 MDM2 抑制剂
简介 肿瘤抑制蛋白 p53 在癌细胞周期中起着至关重要的作用 (1, 2)。大约 50% 的癌症都存在 TP53 基因突变 (2, 3)。在具有 WT p53 的细胞中,由于细胞应激或 DNA 损伤而激活 p53 会导致许多 p53 靶基因的转录激活,从而导致细胞周期停滞、凋亡或衰老 (1, 2, 4)。细胞中的 WT p53 水平受负反馈回路调节。激活的 p53 与 MDM2 基因中的 p53 反应元件结合,导致 MDM2 表达增加。MDM2 蛋白是一种 E3 泛素连接酶,反过来又与 p53 结合并泛素化,导致其被蛋白酶体降解 (5–9)。因此,MDM2 是 p53 的重要调节因子,可以成为具有 WT p53 的癌症的有效治疗靶点。多年来,人们一直对通过药物抑制 MDM2 来稳定 p53 感兴趣,尤其是对于伴有 MDM2 扩增的癌症,包括脂肪肉瘤、尤文氏肉瘤、骨肉瘤和白血病 (2, 10–12)。目前有几种针对 MDM2-p53 相互作用的 MDM2 抑制剂正在临床试验中用于治疗这些癌症 (2),尽管没有一种抑制剂获得 FDA 批准用于任何治疗用途。默克尔细胞癌 (MCC) 是一种高度侵袭性的皮肤神经内分泌癌,发病率很高 (13–15)。MCC 经常转移到淋巴结和远处器官,包括肝脏、骨骼、胰腺、肺和脑 (13–15)。MCC 有两种不同的病因。克隆整合的默克尔细胞多瘤病毒 (MCPyV) 存在于病毒阳性的 MCC (MCCP) 中。这些肿瘤的肿瘤突变负荷较低,具有接近正常的二倍体基因组 (14–20)。相反,病毒阴性 MCC (MCCN) 肿瘤是由慢性紫外线照射引起的,导致高突变负荷和强烈的紫外线突变特征 (14–20)。尽管病因不同,但两种形式的 MCC 都表现出相似的组织学、侵袭性表型和对治疗的反应,表明它们扰乱了相似的致癌途径。虽然 MCCN 通常含有 TP53 和视网膜母细胞瘤肿瘤抑制因子 (RB1) 的功能丧失突变,但 MCCP 通常含有 WT p53 和视网膜母细胞瘤 (RB) 蛋白 (14、15、20–22)。大约 80% 的 MCC 肿瘤是 MCCP,其中大多数具有 WT p53 (16、18、20、23–26)。

MDM2/X抑制剂作为胶质母细胞瘤靶向疗法的放射性抑制剂
抑制MDM2/X-P53相互作用被认为是潜在的抗癌策略,包括治疗胶质母细胞瘤(GB)。 响应细胞应激源(例如DNA损伤),肿瘤抑制蛋白p53被激活,并通过通过DNA修复,细胞周期停滞和凋亡介导细胞损伤来做出反应。 因此,p53激活在细胞存活和癌症疗法的有效性中起着核心作用。 的改变和p53的活性减少发生在25-30%的原发性GB肿瘤中,但该数字在次级GB中急剧增加到60-70%。 因此,建议通过使用靶向分子将突变体p53转换回其野生型形式,或者使用MDM2和MDMX(也称为MDM4)抑制剂将重新激活的p53作为治疗策略。 mdm2降低通过泛素依赖性降解来调节p53活性,并在14%的GB病例中扩增或过表达。 因此,对MDM2的抑制为GB急需新的治疗干预提供了机会。 许多小分子MDM2抑制剂目前正在接受临床评估,无论是单一疗法还是与化学疗法和/或其他靶向药物结合使用。 此外,考虑到p53和MDM2在对辐射诱导的DNA损伤的下游信号反应中的主要作用,MDM2抑制剂与辐射的组合可能为GB治疗提供有价值的治疗性放射敏化方法。 本综述涵盖了MDM2/X在癌症中的作用,在GB中更特定地涵盖了MDM2抑制的潜在放射敏感效应的基本原理。抑制MDM2/X-P53相互作用被认为是潜在的抗癌策略,包括治疗胶质母细胞瘤(GB)。响应细胞应激源(例如DNA损伤),肿瘤抑制蛋白p53被激活,并通过通过DNA修复,细胞周期停滞和凋亡介导细胞损伤来做出反应。因此,p53激活在细胞存活和癌症疗法的有效性中起着核心作用。的改变和p53的活性减少发生在25-30%的原发性GB肿瘤中,但该数字在次级GB中急剧增加到60-70%。因此,建议通过使用靶向分子将突变体p53转换回其野生型形式,或者使用MDM2和MDMX(也称为MDM4)抑制剂将重新激活的p53作为治疗策略。mdm2降低通过泛素依赖性降解来调节p53活性,并在14%的GB病例中扩增或过表达。因此,对MDM2的抑制为GB急需新的治疗干预提供了机会。许多小分子MDM2抑制剂目前正在接受临床评估,无论是单一疗法还是与化学疗法和/或其他靶向药物结合使用。此外,考虑到p53和MDM2在对辐射诱导的DNA损伤的下游信号反应中的主要作用,MDM2抑制剂与辐射的组合可能为GB治疗提供有价值的治疗性放射敏化方法。本综述涵盖了MDM2/X在癌症中的作用,在GB中更特定地涵盖了MDM2抑制的潜在放射敏感效应的基本原理。最后,给出了MDM2/X抑制和p53激活GB的当前状态。

新型小分子 MDM2 抑制剂的设计和计算机模拟研究
摘要:蛋白质-蛋白质相互作用 (PPI) 在许多疾病状况中起着核心作用。因此,靶向和调节 PPI 是治疗疾病的有效方法。癌症也是由于蛋白质-蛋白质相互作用而产生的。在癌症中,肿瘤抑制因子 p53 蛋白被 MDM2 蛋白抑制。p53 蛋白调节细胞周期和细胞凋亡。p53-MDM2 蛋白之间的相互作用导致 p53 功能的抑制。通过这种相互作用,MDM2 降解并抑制 p53 蛋白。因此,靶向和抑制 p53-MDM2 相互作用来治疗癌症是合理的方法。通过用药物靶向这种相互作用,我们可以选择性地杀死癌细胞而不是正常细胞。最近,许多研究人员和制药公司已经报道了 p53-MDM2 相互作用抑制剂药物。并且有几种药物进入了临床试验。在这项研究中,一种新型的 1,2,4-三唑基分子被设计为 MDM2 抑制剂并进行了计算机研究。我们设计了新型化合物 01 和 Lead 1a。在这项工作中,对 Lead 1a 和参考化合物(Nutlin 3a、RG7112)进行了计算机模拟研究。对新型 1,2,4-三唑基 Lead 1a 和参考化合物进行了分子对接研究。发现 Lead 1a 的对接得分优于 Nutlin 3a 并且接近 RG7112。对接研究还确定了各种可能的构象和结合亲和力值。这些结果表明 Lead 1a 是一种潜在的 MDM2 抑制剂和抗癌剂。
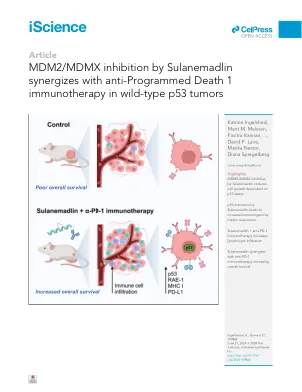
MDM2/MDMX通过sulanemadlin抑制与抗编程死亡1在野生型p53肿瘤中的免疫疗法
摘要免疫疗法已彻底改变了癌症治疗,但其效率取决于肿瘤中强大的免疫反应。沉默抑制肿瘤p53在肿瘤中很常见,可能会影响不同免疫细胞的募集和激活,从而导致免疫逃避和治疗反应不佳。我们发现,p53激活固定肽MDM2/MDMX抑制剂磺胺甲甲酸酯(ALRN-6924)抑制了p53野生型癌细胞在体外和体内的癌细胞生长。在携带p53野生型CT26.WT肿瘤的小鼠中,PD-1抑制剂DX400的单一疗法或磺胺链抑制肿瘤将时间延迟了50%和37%,而联合治疗则将肿瘤倍增时间降低了93%,导致了93%,导致中质生的生存时间增加。sulanemadlin的治疗导致免疫原性和与PD-1抑制作用的联合治疗增加导致淋巴细胞过滤的肿瘤增加。这种组合治疗策略可能会将部分响应者变成免疫疗法的反应者,从而扩大了对PD-1靶向免疫疗法的症状靶标组。

MDM2 抑制联合内分泌治疗和 CDK4/6 抑制治疗 ER 阳性乳腺癌
背景:内分泌治疗耐药是雌激素受体 (ER) 阳性乳腺癌治疗的主要临床挑战。在这种情况下,p53 通常是野生型,其活性可能通过上调其关键调节因子 MDM2 而受到抑制。这构成了我们评估 MDM2 抑制作为治疗耐药性 ER 阳性乳腺癌的治疗策略的理论基础。方法:我们使用 MDM2 抑制剂 NVP-CGM097 单独治疗体外和体内模型以及与氟维司群或哌柏西利联合治疗。我们进行细胞活力、细胞周期、凋亡和衰老测定,以评估 p53 野生型和 p53 突变 ER 阳性细胞系 (MCF-7、ZR75-1、T-47D) 以及对内分泌治疗和 CDK4/6 抑制有耐药性的 MCF-7 系中的抗肿瘤作用。我们进一步评估了药物在内分泌敏感和内分泌抗性的 ER 阳性乳腺癌患者异种移植 (PDX) 模型中的作用。


发现并表征 Brigimadlin,一种适合间歇给药方案的新型高效 MDM2–p53 拮抗剂
p53 被称为基因组的守护者,是最重要的肿瘤抑制因子之一。它在大多数肿瘤中处于失活状态,这是通过肿瘤蛋白 p53 (TP53) 基因突变或关键负调节因子(例如小鼠双微分 2 (MDM2))的拷贝数扩增实现的。与 MDM2 蛋白结合并破坏其与 p53 相互作用的化合物可恢复 p53 肿瘤抑制因子活性,从而促进细胞周期停滞和凋亡。先前使用 MDM2–p53 蛋白–蛋白相互作用拮抗剂 (MDM2–p53 拮抗剂) 的临床经验表明,血小板减少和中性粒细胞减少代表可能限制其治疗效用的靶向剂量限制性毒性。降低给药频率同时保持有效暴露是减轻毒性和改善 MDM2–p53 拮抗剂治疗窗口的一种方法。然而,要实现这一点,需要一种具有优异效力和理想药代动力学特性的分子。在这里,我们介绍了一种新型、在研螺环氧吲哚 MDM2-p53 拮抗剂 brigimadlin (BI 907828) 的发现和表征。Brigimadlin 在临床前模型中表现出高生物利用度和暴露量,以及剂量线性药代动力学。Brigimadlin 治疗恢复了 p53 活性并导致 TP53 野生型、MDM2 扩增癌症临床前模型中的细胞凋亡诱导。以间歇给药方案口服 brigimadlin 在几种 TP53 野生型、MDM2 扩增异种移植模型中诱导了强效的肿瘤生长抑制。探索性临床药代动力学研究 (NCT03449381) 表明,接受口服布吉马林的癌症患者的全身暴露量高,血浆消除半衰期长。这些发现支持继续

MDM2 扩增或过表达的尿路上皮癌患者的致死临床结果以及化疗和免疫治疗耐药性
摘要 背景 E3 泛素连接酶鼠双微体 2 (MDM2) 结合 p53 转录激活结构域并作为 TP53 通路的强效抑制剂,TP53 通路是尿路上皮癌 (UC) 中三种最关键的致癌通路之一。然而,MDM2 扩增在 UC 中的临床意义及其对肿瘤免疫背景的影响仍不清楚。 方法 本研究分析了来自两个当地队列(ZSHS 队列和 FUSCC 队列)的 240 名具有匹配临床注释的 UC 患者。我们通过免疫组织化学分析和靶向测序评估了 MDM2 状态与临床结果、治疗效果和免疫学特征之间的相关性。此外,来自五个独立外部队列的 2264 个 UC 样本(包含基因组、转录组和临床数据)用于验证。结果 MDM2 扩增 (MDM2 Amp) 或蛋白质过表达 (MDM2 OE) 与 UC 患者总体生存率较低 (ZSHS 队列,Log-rank p<0.001;FUSCC 队列,Log-rank p=0.030) 和对铂类化疗 (ZSHS 队列,Log-rank p<0.001) 以及抗 PD-1/PD-L1 免疫疗法 (FUSCC 队列,Log-rank p=0.016) 的反应降低有关,无论 TP53/p53 状态如何。MDM2 扩增或过表达进一步与具有去分化形态的高级别 UC 肿瘤相关。此外,MDM2 扩增或过表达的 UC 与免疫逃避结构相关,其特征是三级淋巴结构浸润比例较低、CD8 + T 细胞、IFN-γ + 细胞、GZMB + 细胞丰度较低,以及免疫检查点分子表达降低,包括程序性死亡配体 1 (PD-L1)、程序性死亡-1 (PD-1) 和细胞毒性 T 淋巴细胞相关蛋白 4 (CTLA-4)。结论 MDM2 扩增或过表达定义了一组致命的 UC 患者,无论 TP53 /p53 状态如何,其预后较差,并且对铂类化疗和免疫疗法均有耐药性。这些肿瘤的特点是去分化形态和免疫抑制微环境。准确

检查更新MDM2抑制剂NAVTEMADLIN阻止了小鼠黑色素瘤的体内生长,并增强放射疗法
1微生物学系,肿瘤和细胞生物学系,瑞典斯德哥尔摩的Karolinska Institutet。 2瑞典乌普萨拉大学免疫学,遗传学和病理学系。 3瑞典乌普萨拉大学乌普萨拉大学外科科学系。 4医学系风湿病学系,卡罗林斯卡大学医院和瑞典斯德哥尔摩的卡罗林斯卡研究所。 5分子医学中心,斯德哥尔摩,瑞典。 6免疫学和过敏部,医学系Solna,Karolinska大学医院和瑞典斯德哥尔摩的Karolinska Institutet。 7 Vanadis Diagnostics,Perkinelmer Inc,Sollentuna,瑞典。1微生物学系,肿瘤和细胞生物学系,瑞典斯德哥尔摩的Karolinska Institutet。2瑞典乌普萨拉大学免疫学,遗传学和病理学系。3瑞典乌普萨拉大学乌普萨拉大学外科科学系。4医学系风湿病学系,卡罗林斯卡大学医院和瑞典斯德哥尔摩的卡罗林斯卡研究所。5分子医学中心,斯德哥尔摩,瑞典。6免疫学和过敏部,医学系Solna,Karolinska大学医院和瑞典斯德哥尔摩的Karolinska Institutet。 7 Vanadis Diagnostics,Perkinelmer Inc,Sollentuna,瑞典。6免疫学和过敏部,医学系Solna,Karolinska大学医院和瑞典斯德哥尔摩的Karolinska Institutet。7 Vanadis Diagnostics,Perkinelmer Inc,Sollentuna,瑞典。

DNA损伤反应抑制剂中的胆管癌
1美国德克萨斯大学医学硕士安德森癌症中心,美国德克萨斯州休斯顿,美国2号,美国2克鲁肯贝格癌症中心,哈雷大学医院,哈雷,德国33790,德国3伊卡尼山上10029年伊卡恩医学院美国纳什维尔37232,美国7弗雷德·哈钦森癌症研究中心,西雅图,西雅图,西雅图98109,美国8血液学系,奥波尔大学,奥波尔大学,45-040,波兰9,波兰9,血液学,肿瘤学,血液质学和干细胞肿瘤学和干细胞植入术(医学院)维多利亚大学医院,M´Alaga,29010,西班牙11 Josa Andr´as教学医院,纽约雷吉,4400,匈牙利12 Kartos Therapeutics,Inc.,Redwood City,CA 94065,美国13 Saint-Louis Hospital&Universit'e Paris´e,Paris”,Paris,75010101010101010101010101010101010年作者 *:+1 713 745 3429; sverstov@mdanderson.org