XiaoMi-AI文件搜索系统
World File Search SystemMyc

先前的免疫力限制了突破性感染期间SARS-COV-2和FC轮廓的粘膜抗体识别
线粒体功能的增加可能会使一些癌症容易受到线粒体抑制剂的影响。由于线粒体DNA拷贝数(mTDNACN)部分调节线粒体功能,因此MTDNACN的准确测量可以帮助揭示哪些癌症是由线粒体功能增加的驱动,并且可能是候选线粒体抑制的候选者。然而,先前的研究采用了巨大的宏观解剖,无法说明MTDNACN中细胞型特异性或肿瘤细胞异质性。这些研究经常产生不清楚的结果,尤其是在前列腺癌中。本文中,我们开发了一种多重原位方法,用于量化细胞类型 - 特异性mtDNACN。我们表明,在前列腺腺癌(PCA)中,高级前列腺上皮内肿瘤(HGPIN)的腔细胞中有MTDNACN增加,并且在转移性castat依 - 耐药的前列腺癌中进一步升高。通过2种正交方法验证了PCA MTDNACN的增加,并伴随着MTRNA和酶活性的增加。从机械上讲,前列腺癌细胞中的MYC抑制作用降低了MTDNA复制和几个mtDNA复制基因的表达,而小鼠前列腺中的MYC激活会导致肿瘤前列腺细胞中的mtDNA水平升高。我们的原位方法还显示,胰腺和结肠/直肠的癌性病变中的mtdnaCn升高,使用临床组织样品表明了对癌症类型的概括。
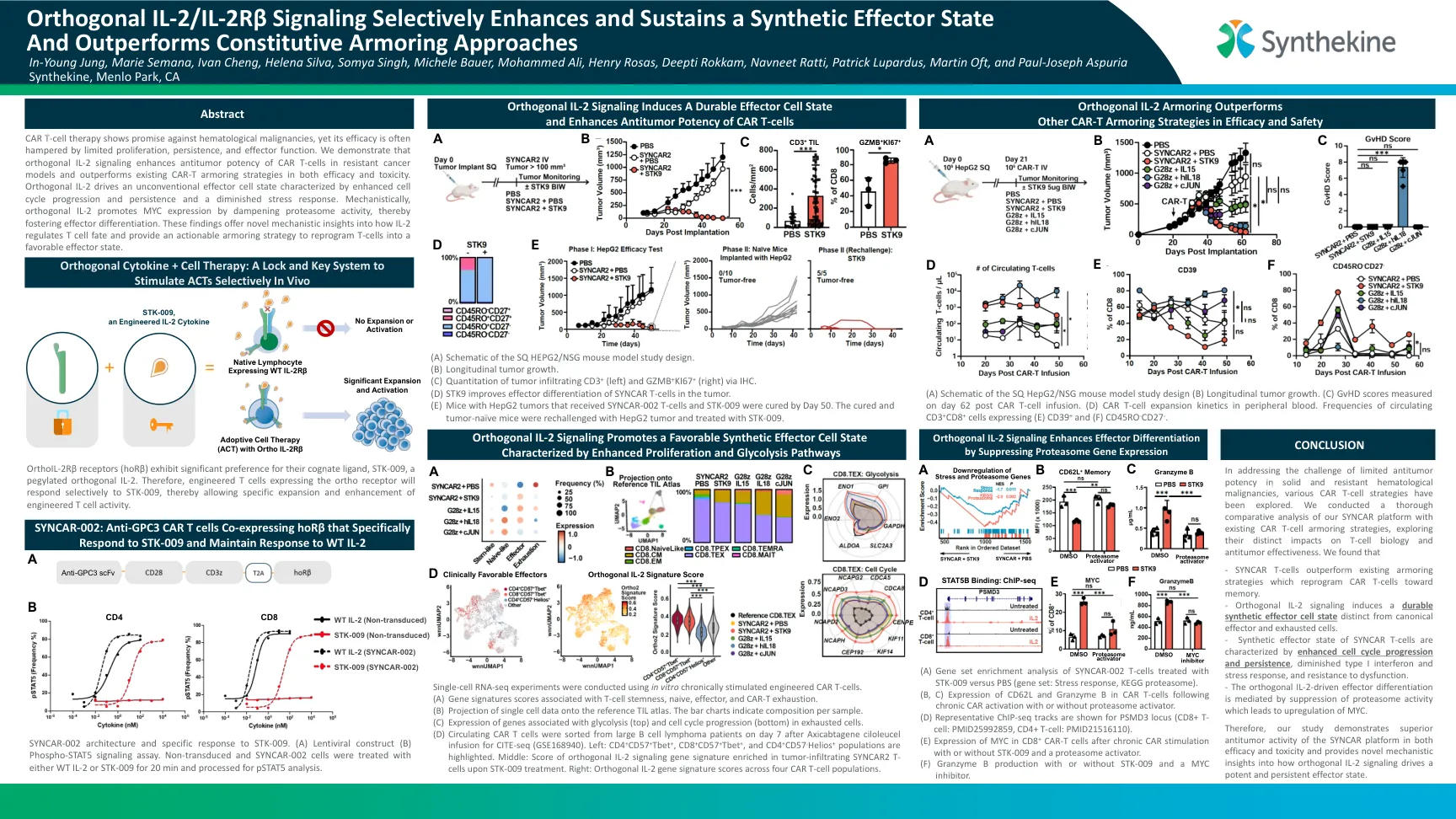
正交IL-2/IL-2Rβ信号有选择地增强和维持合成效应子状态,并且表现优于组成型装甲方法
CAR T细胞疗法表现出对血液学恶性肿瘤的有望,但其功效通常受到有限的增殖,持久性和效应子功能的阻碍。我们证明,正交IL-2信号传导在抗性癌症模型中增强了CAR T细胞的抗肿瘤效力,并且在功效和毒性中都胜过现有的CAR-T装甲策略。正交IL-2驱动非常规效应细胞态,其特征是细胞周期进展和持久性增强以及应力反应减少。 从机械上讲,正交IL-2通过抑制蛋白酶体活性促进MYC的表达,从而促进效应子分化。 这些发现提供了有关IL-2如何调节T细胞命运的新型机械见解,并提供了可行的装甲策略,以将T细胞重编程为有利的效应子状态。正交IL-2驱动非常规效应细胞态,其特征是细胞周期进展和持久性增强以及应力反应减少。从机械上讲,正交IL-2通过抑制蛋白酶体活性促进MYC的表达,从而促进效应子分化。这些发现提供了有关IL-2如何调节T细胞命运的新型机械见解,并提供了可行的装甲策略,以将T细胞重编程为有利的效应子状态。

阿米莉亚项目
乳腺癌通常根据激素受体的表达情况分为亚型,而不考虑其他基因的克隆扩增。我们的实验室发现 14% 的乳腺癌患者有两种致癌基因热休克因子 1 (HSF1) 和细胞性粒细胞瘤病 (c-MYC) 的拷贝数扩增,这两种基因编码同名的转录因子。已知这些转录因子通过多种机制驱动癌症,包括增加干细胞、代谢和存活率。由于转录因子很难用药物靶向,我们旨在用染色质修饰剂的抑制剂间接抑制这些转录因子的活性。我们使用卵巢癌细胞系作为模型进行了药物筛选,以找到染色质修饰剂抑制剂,这些抑制剂可以选择性地杀死 HSF1 和 c-MYC 基因扩增的细胞。在药物筛选中,我们发现携带 HSF1- MYC 共扩增的细胞系对组蛋白去乙酰化酶 (HDAC) 抑制剂的敏感性高于没有这种共扩增的癌细胞系。我们的初步结果表明,HDAC 抑制剂 Entinostat 可降低 HSF1 和 c-MYC 蛋白水平,并降低其直接下游靶标的 mRNA 表达。我们的实验室将进一步研究组蛋白去乙酰化酶抑制剂 Entinostat 在含有 HSF1 和 c-MYC 扩增的癌症中降低 HSF1 和 c-MYC 的机制,以及 Entinostat 在这些癌症中的疗效。

利用 hei-tag 促进靶向基因组编辑
摘要 CRISPR/Cas9 的精确靶向基因组编辑是模型和非模型系统中基础研究和转化方法的关键。尽管迄今为止在所有测试的物种中都处于活跃状态,但编辑效率仍有改进空间。细菌 Cas9 需要通过与核定位信号 (NLS) 融合有效地穿梭到细胞核中。通常会添加额外的肽标签(例如 FLAG 或 myc 标签)以立即检测或直接纯化。通常通过施用预组装的蛋白质/RNA 复合物来获得即时活性。我们提出了“hei 标签(高效标签)”,它可以在以 mRNA 形式提供时增强 CRISPR/Cas 基因组编辑工具的活性。将 hei 标签(一种通过灵活的接头与优化的 NLS 偶联的 myc 标签)添加到 Cas9 或 C-to-T(胞嘧啶到胸腺嘧啶)碱基编辑器中可显著提高各自的靶向效率。这导致双等位基因编辑增加,但等位基因变异减少,表明即使在早期发育阶段也具有即时活性。hei-tag boost 在从鱼类到哺乳动物的模型系统中都很活跃,包括组织培养应用。只需简单添加 hei-tag,即可立即升级现有且可能高度适应的系统,并建立可立即应用于 mRNA 水平的新型高效工具。

免疫破坏是否会驱动所有形式的骨髓衰竭?
化学疗法或内源性醛。ICL的形成触发FA核心综合以定位于DNA病变,然后募集其他含FA蛋白质的复合物和ICL修复酶(4)。FA基因中的不活性突变导致无法修复ICL,从而导致杂种不稳定性。 FA造血干细胞和祖细胞(HSPC)中的DNA损伤会激活MYC和遗传毒性应激/TP53途径,并诱导炎症的炎性细胞因子信号传导(1,5,6)。 通过先前未定义的下流事件,这些变化驱动了FA中的HSPC损失和/或血液系统恶性肿瘤。 在JCI的这个问题中,Casado等。 通过识别免疫介导的机制来提供这种缺失的下游链接,通过该机制激活DNA损伤途径会导致BMF(7)(图1)。FA基因中的不活性突变导致无法修复ICL,从而导致杂种不稳定性。FA造血干细胞和祖细胞(HSPC)中的DNA损伤会激活MYC和遗传毒性应激/TP53途径,并诱导炎症的炎性细胞因子信号传导(1,5,6)。通过先前未定义的下流事件,这些变化驱动了FA中的HSPC损失和/或血液系统恶性肿瘤。在JCI的这个问题中,Casado等。通过识别免疫介导的机制来提供这种缺失的下游链接,通过该机制激活DNA损伤途径会导致BMF(7)(图1)。

患者问题:肺癌的创新
ALK (4) NRAS (2) ARID1A (1) NRG1 (2) ATM (2) NTRK (6) ATR (1) P53 (1) BRAF (15) PALB (1) BRCA (2) PDGFR (2) CCND (1) PIK3 (5) CDK4/6 (1) POLD1 (2) CDKN2 (1) POLE (2) ctDNA (3) PRKC (2) CTNNB1 (1) PTCH1 (2) DDR (5) RAF (2) EGFR (10) RAS (2) ERK (2) RET (4) EWSR1 (1) ROS1 (5) EZH2 (1) SMO (1) FBXW7 (1) STK11 (1) FGFR (4) TAF15 (1) GNAQ/11 (2) TMB (4) HRD (7) TP53 (1) IDH (2) TSC (1) KIT (2) UGT1A1 (1) KRAS (24) CCNE1 (1) MDM2 (1) CLDN (3) MEK (3) FET (1) MET (5) FRa (1) MMR (2) FUS (1) MRD (2) HER2 (12) MSI-H (1) MAGE (2) MTAP (4) MUC (1) MYC (1) PD-L1 (12) Nectin-4 (2) PRAME (1) NF1 (6)

细胞疗法
最受过临床试验的细胞,间充质基质细胞(MSC)现在已知主要通过包括外泌体的旁分泌分泌发挥其治疗活性。为了减轻对MSC外泌体制备的可伸缩性和可重复性的潜在调节问题,使用高度表征的MYC降低的单克隆细胞系产生MSC外泌体。这些细胞不会在无胸腺裸鼠或表现出与锚固无关的生长中形成肿瘤,并且它们的外泌体不携带MYC蛋白或促进肿瘤生长。与腹膜内注射不同,MSC在IMQ诱导的牛皮癣的小鼠模型中的局部应用减轻了白介素(IL)-17,IL-23和末端组合复合物,C5B9在牛皮乳肌皮中。应用于人类皮肤外植体时,从共价标记的荧光MSC外泌体的荧光渗透并持续在角质层中,大约24小时,而从角质层中忽略不计,将其从角膜层中忽略不计。作为牛皮癣的角膜层的特征在于活化的补充和Munro微鳞片,我们假设局部施用的外泌体渗透到牛皮癣的角膜层以抑制C5B9补体通过CD59抑制CD59,并且这种抑制作用抑制了中性粒细胞粒细胞的IL IL-17。与此相一致,我们证明了C5B9在纯化的人类嗜中性粒细胞诱导的IL-17分泌上的组装,MSC外泌体使这种诱导构成了这种诱导,这又被中和中和的抗CD 59抗体所消除。因此,我们确定了通过局部应用外泌体缓解银屑病IL-17的作用机理。由Elsevier Inc.出版©2023国际细胞和基因治疗学会。这是CC BY-NC-ND许可(http://creativecommons.org/licenses/by-nc-nd/4.0/)下的开放访问文章

Monjuvi(Tafasitamab-cxix)
概述此文档解决了Monjuvi(Tafasitamab-cxix)的使用。Monjuvi是一种CD19指导的细胞溶剂抗体FDA,与周期性胺相结合,用于治疗成年患者的复发或难治性弥漫性大B细胞淋巴瘤(DLBCL),包括未指定的DLBCL,包括由低级淋巴瘤和较低级别的pollyology Comply compl and Olig andology Compliention(包括DLBCL),以及(NCCN)提供了其他建议,并提供了2A类使用Monjuvi的证据。建议与Lenalidomide联合治疗组织学转化以扩散大型B细胞淋巴瘤(DLBCL),而不会在不候选移植并接受过

赖氨酸和蛋氨酸对牛乳腺上皮细胞候选转录因子mRNA表达的影响
已确定必需氨基酸 (EAA) 通过快速改变翻译因子的磷酸化状态来调节乳腺上皮细胞的蛋白质合成。然而,对 EAA 供应的长期转录反应研究得很少。选定了八种转录因子作为 EAA 通过氨基酸反应 (ATF4、ATF6)、丝裂原活化蛋白激酶 (JUN、FOS、EGR1) 和雷帕霉素复合物 1 的机制靶点 (MYC、HIF1A、SREBF1) 影响乳腺细胞功能的候选介质。目的是确定在施加 EAA 缺乏 24 小时后,这些候选基因的表达是否以及何时在牛乳腺上皮细胞原代培养物中受到影响,并评估 EAA 缺乏对蛋白质合成、内质网大小、细胞增殖和脂肪形成的影响。将分化细胞在代表所有氨基酸的正常生理浓度 (CTL)、低赖氨酸 (LK) 或低蛋氨酸 (LM) 的 3 种处理培养基中的 1 种中培养 24、40、48 或 60 小时。LK 和 LM 均抑制蛋白质合成并激活 ATF4 表达,表明经典的氨基酸反应途径已被触发。然而,LK 或 LM 对内质网大小没有影响,可能与 LM 上 ATF6 表达升高有关。早期反应基因 JUN 、 FOS 、 EGR1 和 MYC 的表达没有因 EAA 缺乏而升高,但 LM 降低了 EGR1 的表达。LM 还增加了 HIF1A 的表达。EGR1 和 HIF1A 的表达结果与观察到的细胞增殖率下降一致。不同时间点 SREBF1 表达对 LK 和 LM 的不同反应可能导致对脂肪生成率没有影响。这些发现表明,EAA 缺乏可能通过转录因子抑制乳腺蛋白质的合成和细胞增殖。
