XiaoMi-AI文件搜索系统
World File Search SystemPromoter

胃肠道肿瘤免疫中的表观遗传启动子改变
摘要 目的 癌症的表观基因组改变与免疫微环境相互作用,决定肿瘤的发展和治疗反应。我们旨在研究胃癌中表观遗传替代启动子使用对肿瘤免疫微环境的调节,并将我们的研究结果扩展到其他胃肠道肿瘤。设计 使用一种新颖的生物信息学算法 (proActiv) 量化替代启动子负荷 (APB),以从短读 RNA 测序和分为 APB 高、APB int 和 APB 低的样本推断启动子活性。进行单细胞 RNA 测序以分析肿瘤内免疫微环境。人源化小鼠癌症体内模型用于探索肿瘤动力学、替代启动子使用和人体免疫系统之间的动态时间相互作用。评估了接受免疫疗法治疗的多组胃肠道肿瘤,以确定 APB 与治疗结果之间的相关性。结果 APB high 胃癌肿瘤表达的 T 细胞溶细胞活性水平降低,并表现出免疫耗竭的特征。单细胞 RNA 测序分析证实了 APB high 肿瘤中不同的免疫群体和较低的 T 细胞比例。使用具有活跃人类免疫系统的“人源化小鼠”进行的功能性体内研究揭示了 APB 与肿瘤生长之间的明显时间关系,其中 APB high 肿瘤几乎没有人类 T 细胞浸润。对接受免疫疗法治疗的胃肠道癌患者的分析证实了 APB high 肿瘤对免疫检查点抑制的耐药性。与 APB low 相比,APB high 胃癌的无进展生存期明显较差(中位数 55 天 vs 121 天,HR 0.40,95% CI 0.18 至 0.93,p=0.032)。结论这些发现表明替代启动子的使用与肿瘤微环境之间存在关联,从而导致免疫逃避和免疫疗法耐药性。

启动子的全基因组核小体分辨率图
摘要 增强子-启动子环路模型长期以来一直主导着基因调控领域,其中增强子通过物理接触激活其靶基因。然而,由于存在替代机制的证据以及缺乏系统验证(主要是由于缺乏合适的实验技术),该模型的普遍性受到了质疑。在本研究中,我们提出了一种新的基于 MNase 的邻近连接方法,称为 MChIP-C,该方法可以在基因组范围内以单核小体分辨率测量蛋白质介导的染色质相互作用。通过应用 MChIP-C 研究 K562 细胞中以 H3K4me3 启动子为中心的相互作用,我们发现与基于限制性内切酶的 C 方法相比,它具有大大提高的分辨率和灵敏度。这使我们能够将 EP300 组蛋白乙酰转移酶和 SWI/SNF 重塑复合物确定为建立和/或维持增强子-启动子相互作用的潜在候选者。最后,利用已发表的 CRISPRi 筛选数据,我们发现大多数经过功能验证的增强子确实与其同源启动子发生物理相互作用,支持增强子-启动子环路模型。

接下来会发生粘结启动子树脂...
Resin Developed Pre-Commercial Designation Year Developed Compostion Penacolite® B1A 1940's Resorcinol - Formaldehyde Penacolite® B16 1950's Resorcinol - Formaldehyde Penacolite® B17 1962 Resorcinol - Formaldehyde Penacolite® B18 1964 Resorcinol - Formaldehyde Penacolite® B18S RDL - 065 1976 RF Resin + RM441 Penacolite® B19S RDL - 095 1978 RF Resin + RM441 Penacolite® B20S*** RDL - 055 1988 R + Styrene + F Resin Penacolite® B21S *** RDL - 516 2004 R + Styrene + F Resin Penacolite® B22 1950's Phenol + Resorcinol + FPenaColite®untipro 100 ***良好的年份代码:“ Rajic” 2003 M-羟基二苯胺Penacolite®I868 C *** CRL -434 2009 RESORCINOL -MDI ADRAUCT -MDI ADRAUCT

真空镀铝 CPP 薄膜用金属附着力促进剂
自从柔性薄膜发明以来,包装行业的发展达到了惊人的高度。通过制造实践对这些薄膜的利用不断创新,对最适合进一步增强其理想特性的广泛产品的需求不断增长。虽然柔性包装行业使用了许多聚合物,但最常见的是聚丙烯 (PP)、聚乙烯 (PE)、聚氯乙烯 (PVC)、聚对苯二甲酸乙二醇酯 (PET)。最近发生的最受欢迎的创新之一是将金属化薄膜引入柔性包装行业。通过在真空条件下在聚合物表面沉积一层极薄的铝蒸气,成品被证明具有更好的防水和防氧性能,并提供金属光泽的外观——非常像铝箔。能够提供铝箔所具有的一些关键特性,同时成本和皮重却低得多,这使得金属化薄膜成为食品和糖果领域的热门选择。聚酯和 PP 薄膜是生产这些高阻隔包装薄膜(金属化薄膜)最常用的基材。铸造 PP 薄膜正迅速成为包装行业首选的金属化基材。真空金属化铸造 PP 薄膜在行业中被称为 VMCPP 或简称为 MCPP。应用领域包括食品包装、化妆品包装、烟草和酒类包装,以及众多装饰应用。MCPP 薄膜的阻隔性能取决于薄膜表面金属附着力的质量,而这又取决于:

酵母 PHO5 启动子处核小体结构的有效动力学
摘要染色质动力学由重塑酶介导,在基因调控中起着至关重要的作用,正如在典型模型酿酒酵母 PHO5 启动子中建立的那样。然而,有效的核小体动力学,即启动子核小体配置的轨迹,仍然难以捉摸。在这里,我们通过整合已发表的单分子数据推断出这种动力学,这些数据捕获了从受抑制到完全活跃的 PHO5 启动子状态的多核小体配置,以及其他现有的组蛋白周转和新的染色质可及性数据。我们设计并系统地研究了一类新的“受调节的开关滑动”模型,模拟全局和局部核小体(解)组装和滑动。68,145 个模型中只有 7 个与所有数据吻合良好。所有七个模型都涉及滑动和 N-2 核小体的已知核心作用,但通过调节一个组装而不是解体过程来调节启动子状态转换。这与 PHO5 启动子先前观察结果的常见解释一致,但提出了挑战,并表明染色质通过结合竞争而开放。

胶质瘤干细胞作为胶质瘤进展的促进剂 - IRIS
摘要:神经胶质瘤的侵袭性和对治疗的抵抗性使其成为肿瘤学的一个主要问题。尽管医学科学取得了重大进步,但神经胶质瘤的预后仍然不容乐观,手术、放疗 (RT) 和化疗 (CT) 等传统治疗方法经常被证明无效。在发现神经胶质瘤干细胞 (GSC) 后,将神经胶质瘤视为均质肿块的传统观点发生了变化。GSC 对肿瘤生长、治疗抵抗和复发至关重要。这些细胞独特的分化和自我更新能力正在改变我们对神经胶质瘤生物学的认识。本系统文献综述旨在揭示与 GSC 相关的神经胶质瘤进展的分子驱动机制。系统综述遵循 PRISMA 指南,在 PubMed、Ovid MED-LINE 和 Ovid EMBASE 上进行了彻底的文献检索。第一次文献检索于 2024 年 3 月 1 日进行,搜索更新于 2024 年 5 月 15 日。搜索使用 MeSH 术语和布尔运算符,重点关注与 GCS 介导的胶质瘤进展相关的分子机制。纳入标准包括英文研究、临床前研究和临床试验。最初确定了 957 篇论文,其中 65 篇从 2005 年到 2024 年的研究最终被纳入审查。主要 GSC 模型分布按频率降序排列:U87:20 项研究(32.0%);U251:13 项研究(20.0%);A172:4 项研究(6.2%);和 T98G:2 项研究(3.17%)。从最频繁到最不频繁,主要 GSC 通路的分布如下:Notch:8 项研究(12.3%);STAT3:6 项研究(9.2%); Wnt/β-catenin:6 项研究(9.2%);HIF:5 项研究(7.7%);PI3K/AKT:4 项研究(6.2%)。分子效应的分布(从最常见到最不常见)如下:抑制分化:22 项研究(33.8%);增加增殖:18 项研究(27.7%);增强侵袭能力:15 项研究(23.1%);增加自我更新:5 项研究(7.7%);抑制细胞凋亡:3 项研究(4.6%)。这项研究突出了 GSC 异质性和胶质母细胞瘤微环境中的动态相互作用,强调需要采取量身定制的方法。影响 GSC 行为的一些关键通路是 JAK/STAT3、PI3K/AKT、Wnt/β-catenin 和 Notch。治疗可以针对这些通路。这项研究敦促进行更多研究以填补 GSC 生物学方面的知识空白,并将研究结果转化为有用的治疗方法,以改善 GBM 患者的治疗结果。

通过 CRISPR/Cas9 创建的天然启动子-基因融合……
本文已接受出版并经过完整的同行评审,但尚未经过文字编辑、排版、分页和校对过程,这可能会导致此版本与记录版本之间存在差异。请引用本文 doi:10.1002/1873-3468.14365
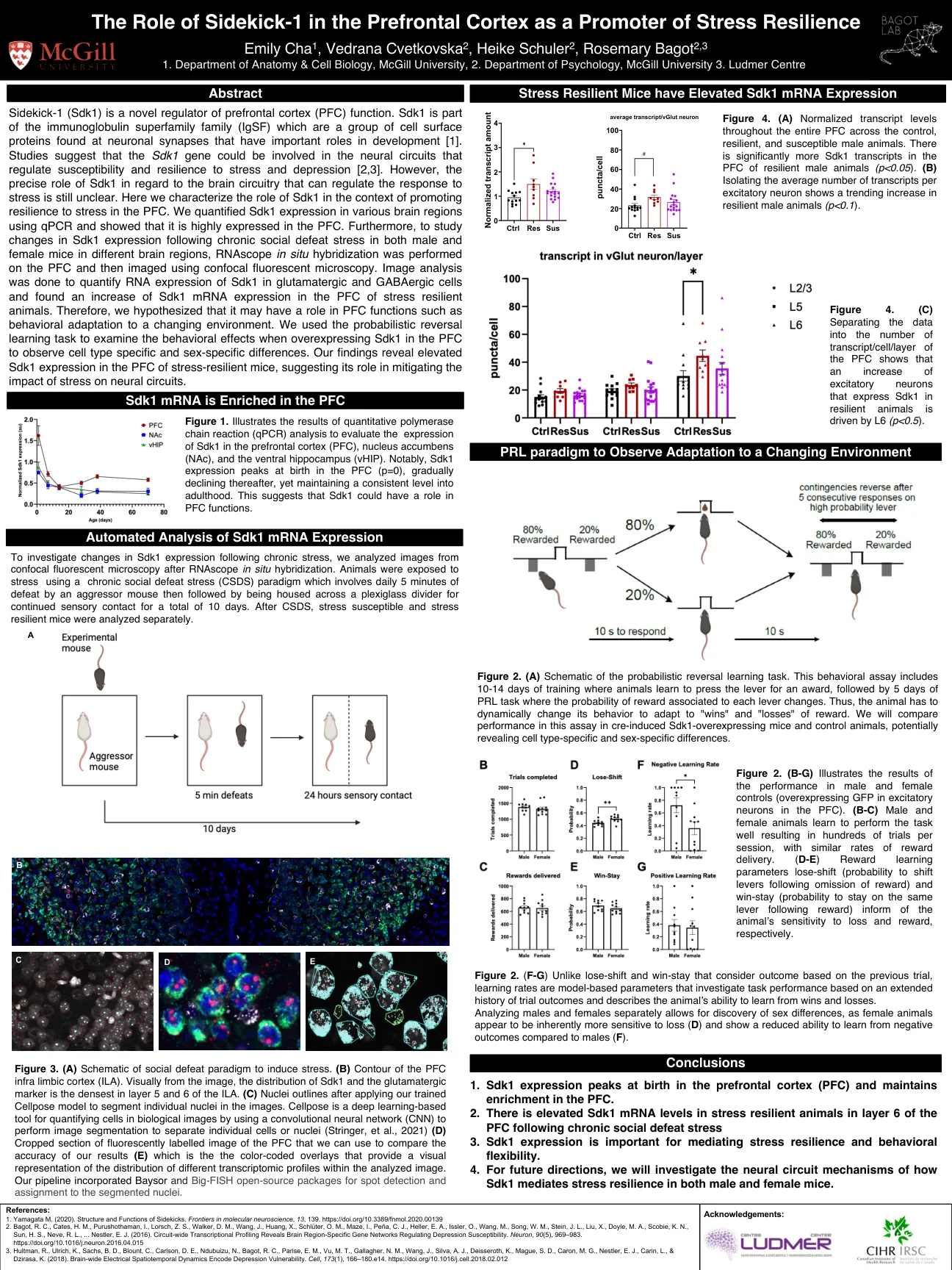
sidekick-1在前额叶皮层中的作用作为应力弹性的启动子
sidekick-1(SDK1)是前额叶皮层(PFC)功能的新型调节剂。SDK1是免疫球蛋白超家族(IGSF)的一部分,它们是在神经元突触中发现的一组细胞表面蛋白,它们在发育中具有重要作用[1]。研究表明,SDK1基因可能参与调节压力和抑郁症的易感性和韧性的神经回路[2,3]。但是,SDK1在可以调节应力反应的脑电路方面的确切作用尚不清楚。在这里,我们表征了SDK1在促进PFC压力的韧性中的作用。我们使用qPCR量化了各个大脑区域中的SDK1表达,并表明它在PFC中高度表达。此外,要研究不同大脑区域的男性和雌性小鼠慢性社交失败压力后SDK1表达的变化,在PFC上进行了原位杂交,然后使用共斑荧光显微镜进行成像。进行图像分析以量化谷氨酸能和GABA能细胞中SDK1的RNA表达,并发现在应激弹性动物的PFC中发现SDK1 mRNA表达增加。因此,我们假设它可能在PFC函数中起作用,例如行为适应不断变化的环境。我们使用概率逆转学习任务来检查PFC中SDK1过表达的行为效应,以观察特定的细胞类型和性别特异性差异。我们的发现显示在应激势力小鼠的PFC中SDK1的表达升高,这表明其在减轻压力对神经回路的影响中的作用。

番茄果实外果皮优先启动子的鉴定及在遗传工程中的应用
番茄 ( Solanum lycopersicum ) 是一种全球性种植的作物,具有巨大的经济价值。外果皮决定了番茄果实的外观,并在收获前和收获后保护其免受各种生物和非生物挑战。然而,目前还没有番茄外果皮特异性启动子,这阻碍了基于外果皮的基因工程。在这里,我们通过 RNA 测序和逆转录-定量 PCR 分析发现番茄基因 SlPR10 ( PATHOGENESIS RELATED 10 ) 在外果皮中大量表达。由 2087-bp SlPR10 启动子 ( pSlPR10 ) 表达的荧光报告基因主要在 Ailsa Craig 和 Micro-Tom 品种的转基因番茄植株的外果皮中检测到。该启动子进一步用于番茄中 SlANT1 和 SlMYB31 的转基因表达,它们分别是花青素和角质层蜡质生物合成的主要调节因子。pSlPR10 驱动的 SlANT1 表达导致花青素在外果皮中积累,赋予果实抗灰霉病和延长保质期,而 SlMYB31 表达导致果皮蜡质增厚,延缓水分流失并延长果实保质期。有趣的是,pSlPR10 和另外两个较弱的番茄外果皮优先启动子在转基因拟南芥 (Arabidopsis thaliana) 植物的子房中表现出一致的表达特异性,这不仅为番茄外果皮和拟南芥子房之间的进化同源性提供了线索,而且为研究拟南芥子房生物学提供了有用的启动子。总的来说,这项研究报告了一种理想的启动子,能够在番茄外果皮和拟南芥雌蕊中实现靶基因表达,并证明了其在番茄果实品质遗传改良中的实用性。

乳酸:肝癌微环境促进剂的叙述性综述
肿瘤诱导的免疫细胞募集是对早期疾病的早期反应,通过启动致癌炎症,导致免疫介导促进肿瘤细胞增殖、存活和血管生成 (5)。如果早期流入的免疫细胞成功检测到异常并发起免疫反应,恶性细胞将被直接消灭。但是,如果信号或反应不足导致免疫逃避,肿瘤就会发展并开始局部生长,最终扩散到远处。肿瘤的发展伴随着一系列可溶性因子,这些因子促进非恶性细胞、血管和基质的流入,共同形成肿瘤微环境 (TME) (6)。随着肿瘤的进展,TME 逐渐发生变化,可能变得非常复杂。因此,对 TME 本质的研究有望带来改善治疗反应的新干预措施。