XiaoMi-AI文件搜索系统
World File Search SystemSecrete

肿瘤衍生的小细胞囊泡充当治疗性纳米粒子递送的障碍
纳米颗粒有望用于药物输送应用,并具有多种临床批准的产品。但是,在实体瘤中获得高纳米颗粒的积累仍然具有挑战性。在这里我们表明,肿瘤细胞衍生的小细胞外囊泡(SEV)将纳米颗粒递送到肿瘤,揭示了基于纳米颗粒的肿瘤疗法的另一个障碍。肿瘤细胞在肿瘤微环境中分泌大量的SEV,然后结合进入肿瘤组织的纳米颗粒并将其传递到肝库普弗细胞中以降解。敲低Rab27a是一种控制SEV分泌的基因,可降低SEV水平并改善肿瘤组织中纳米颗粒的积累。与Rab27a在脂质纳米颗粒中共同包裹的抑制肿瘤和炎性蛋白质的信使RNA的治疗功效大大提高。一起,我们的结果表明,肿瘤细胞衍生的SEV是针对纳米颗粒肿瘤递送的防御系统,并且该系统可能是改善基于纳米颗粒的肿瘤疗法的潜在靶标。
角质形成细胞感知并通过刺激/IFN-κ激活和APOBEC3G诱导消除CRISPR DNA
引言角质形成细胞(KC)长期以来一直被认为是转染最困难的细胞类型之一(1,2),但是这种耐药性背后的机制尚不清楚。kc是表皮的主要细胞成分,它充当人体和外部药物(例如细菌和病毒)之间的主要界面。除了对上皮真皮作为物理屏障的关键贡献之外,KC还通过一系列模式受体和分泌各种细胞因子的能力具有非常活跃的免疫作用(3)。原核生物衍生的定期插入短的短质体重复序列(CRISPR)/CAS9技术改变了我们操纵活细胞中特定DNA和RNA序列的能力(4)。CRISPR/CAS9系统依赖于引导RNA(GRNA)来靶向特异性和功能,通过产生焦油的DNA断裂,从而通过各种内源性机制刺激修复。CRISPR技术可用于通过非同源最终连接途径或通过同源性编辑的单基结合途径插入或删除小型DNA段 -

IL-17 and-il-22-as-as-as-stable-...
IL-17和IL-22之间的相互作用对于维持组织稳态至关重要[9]。在发现Th17细胞作为T-Helper细胞的独立线[10,11]后,它们在炎症中的作用变得更加明显,它分泌IL-17和IL-22细胞因子。仍然需要澄清其分泌的刺激,但是在人类中,与小鼠模型不同,它们并不总是协同分泌的[12,13]。通常,IL-17主要是一种炎性细胞因子,通过增强抗菌肽,粒细胞和中性粒细胞在周围组织中积累的上皮上皮释放来预防感染,而IL-22则是一种稳态的稳态,仅是一种固定的稳态,仅是造成边界组织和组织的整体性启动性的,并且是造成边界组织和组织的构成的,并且是构成的,并且是构成的,并且是构成的,并且是构成的,并且是一种稳固的细胞影响。 [13,14]。IL-17和IL-22被认为是组织信号细胞因子,因为它们主要影响固体组织的细胞[12,15,16]。其受体的分布大大分歧。在人类中,IL-17的受体复合物在上皮,间质和造血细胞上广泛表达[13,17]。il-17和Th17细胞与各种器官(例如肝脏或皮肤)的促炎和纤维化条件有关[18,19]。

CD8+ T细胞策划的T细胞免疫
CD8 + T细胞属于自适应免疫系统在防御病毒感染和癌症中起关键作用。当前的基于CD8 + T细胞的免疫疗法已成为消除肿瘤细胞和长期预防血液系统恶性肿瘤复发的优质治疗途径。据信,有效的适应性免疫反应主要依赖于先天室的帮助。不变的天然杀手T(inkt)细胞是先天性的T淋巴细胞,被认为是首先对感染反应的一些细胞,可以分泌大量不同的细胞因子和趋化因子,以广泛调节先天和适应性免疫应答者。像CD8 + T细胞一样,Inkt细胞在防御细胞内致病感染和癌症中也起着重要作用。在这篇综述中,我们将讨论Inkt细胞贡献的CD8 + T细胞免疫,包括inkt细胞介导的交叉染色和记忆形成,并讨论我们对我们对内存记忆CD8 + T细胞差异机制的理解的最新进展,以及T细胞免疫的衰老受损。

T辅助细胞分化和... 的分子机制T辅助细胞分化和...
病原体可以感染许多不同的组织和细胞室,这些病原体的生命周期可能会有很大的变化。为了克服这一点,宿主免疫系统已经制定了病原体特异性策略。例如,TH细胞(效应CD4 T细胞)针对许多不同的病原体协调免疫反应(1-4)。幼稚的CD4 T细胞通过血液和淋巴系统循环,通过与淋巴组织中树突状细胞(DC)呈现的抗原结合来收集有关病原体的信息。dcs在将这些病原体特异性抗原呈现给幼稚的CD4 T细胞时,还会分泌细胞因子,从而触发分化途径(1-4)。活化的CD4 T细胞增殖并分化为不同的免疫细胞亚群,这些亚群确定了免疫反应的类型(1-4)。在大约30年前,在细胞分化和功能领域的研究开始了,但仍在进行中。在这篇评论中,我将总结控制细胞分化的基本原理(图1)。该领域的其他主题,包括微生物群和可塑性/异质性,将在本期其他文章中涵盖。

微生物群对牙周组织炎症性疾病发生发展的影响
口腔器官和组织功能的一个重要特征是,其中发生的所有过程都是在各种微生物的不断存在下进行的,这些微生物会导致体内病理过程的发展或与之相关。在慢性全身性牙周炎的发病机制中,牙菌斑穿透牙龈沟底部,渗透到上皮下方进入结缔组织基质,引起其炎症。细菌会产生多种对周围组织有毒性作用的毒性物质。大多数细菌会产生链状脂肪酸,抑制白细胞和吞噬细胞的趋化作用。厌氧菌和螺旋体会分泌多种对大多数组织具有极大毒性的物质(丙酸和吲哚)。牙周组织中的炎症是由牙菌斑生物膜的微生物群引起的。随着牙周炎的发展,牙龈卟啉单胞菌、中间卟啉单胞菌和福赛丝菌的数量增加了100多倍。因此,给出的数据证明,在牙周组织炎症过程的发展和过程中,涉及复杂的菌群失调和组织-细胞相互作用,其动态平衡取决于其结果。

荨麻疹和其他肥大细胞疾病的新治疗方法
图 3 肥大细胞疾病治疗靶点。肥大细胞的逐步激活使得治疗靶向激活的每个阶段成为可能:(1) 肥大细胞在与已知可避免的触发因素相互作用时被激活。(2) 肥大细胞的激活是通过 IgE 依赖性 Fc ε RI 激活或 IgE 非依赖性激活受体实现的,这可分别被 IgE 抑制剂和其他激活分子的抑制剂阻断。(3) 激活的肥大细胞分泌脂质介质(例如白三烯、前列腺素、血小板活化因子)、细胞因子、趋化因子和生长因子,(4) 经历含有预先形成的介质(例如组胺、蛋白酶、细胞因子)的分泌颗粒脱颗粒;这些细胞外介质可在分泌后被阻断,以抑制导致炎症的下游过程。 (5) MC 迁移、增殖和存活依赖于 KIT,这是一种 SCF 结合受体,其激活可被 KIT 抑制剂阻断,从而通过 MC 耗竭来抑制 MC。 (6) 通过激动剂配体与抑制性受体(如 Siglec-8)结合来阻断 MC 激活,可用于 MC 沉默策略。
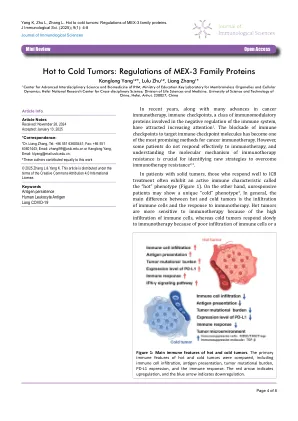
热肿瘤:MEX-3家族蛋白的法规
肿瘤细胞免疫逃生的机制许多机制,一些研究将其定义为“ 3C”模型6,即伪装,它使肿瘤细胞无法通过免疫细胞识别。胁迫,直接或间接干扰免疫细胞;和细胞保护作用,这是对肿瘤细胞免受免疫细胞毒性的保护。这些机制主要包括肿瘤抗原表达或表现中的缺陷:肿瘤细胞可能会失去或改变其表面抗原的表达,从而使T细胞无法识别和攻击它们;免疫抑制细胞的募集:各种免疫抑制细胞(例如调节性T细胞,髓样抑制细胞等)可以募集到肿瘤微环境中,这可以直接抑制效应T细胞的功能。和细胞因子分泌:肿瘤细胞可以分泌一些免疫抑制性细胞因子(例如IL-10,TGF-β等)抑制免疫反应并促进肿瘤的生长;肿瘤微环境的变化:肿瘤细胞可以改变微环境,使其更有利于肿瘤生长并抑制抗肿瘤免疫反应。这些机制经常重叠,从而使肿瘤有效地逃避了免疫系统监测。

单细胞水平的病毒感染与先天免疫之间的对抗
被病毒感染时,细胞可能会分泌干扰素(IFN),该干扰素(IFN)促使附近细胞为即将到来的感染做准备。相互,病毒蛋白通常会干扰IFN合成和IFN诱导的信号传导。我们使用基于药物的随机方法对传播病毒与先天免疫反应之间的串扰进行了建模。通过分析免疫荧光显微镜图像,我们观察到呼吸综合病毒(RSV)(RSV)和感染A549细胞之间的相互拮抗作用会导致单细胞水平和复杂的细胞信号传导状态空间模式的二分法反应。我们的分析表明RSV在三个层面上阻止了先天反应:通过抑制IRF3激活,抑制IFN合成以及抑制STAT1/2激活。反过来,由IFN刺激(STAT1/2激活)基因编码的蛋白质抑制了病毒RNA和病毒蛋白的合成。这些抑制作用的显着结果是病毒蛋白缺乏巧合和单个细胞中IFN的表达。该模型可以研究免疫刺激有缺陷的病毒颗粒和信号网络扰动的影响,这些影响可能有可能促进病毒感染的遏制或清除。

LitterGuard® LT-C
产品描述:LitterGuard LT-C 用于对健康怀孕母猪和母猪进行疫苗接种,以将保护性母体抗体被动转移给猪,有助于预防由 C 型产气荚膜梭菌产生的β毒素和产生热不稳定毒素或具有 K99、K88、987P 或 F41 粘附因子的大肠杆菌肠毒素菌株引起的新生儿腹泻。细菌毒素是由化学灭活的大肠杆菌菌株和 C 型产气荚膜梭菌β毒素制备的。使用无菌佐剂来增强免疫反应。疾病描述:大肠杆菌肠毒素菌株是猪新生儿腹泻最重要的病原体之一。研究表明,从腹泻猪身上分离出的产肠毒素大肠杆菌具有两个共同特征:(1)它们具有菌毛,即细菌附着在肠道上皮细胞上的表面抗原结构;(2)它们表达肠毒素,导致肠道细胞将体液和电解质分泌到肠腔中。结果导致腹泻、脱水,严重时甚至死亡。与猪新生肠大肠杆菌病相关的 4 种主要菌毛类型是 K99、K88、987P、1 和 F41。2