XiaoMi-AI文件搜索系统
World File Search SystemUbiquitination

在26S蛋白酶体调节亚基RPN2基因中,恶性疟原虫赋予青蒿素的抗性
减轻疟疾和相关死亡的负担受到了疟疾寄生虫能够发展对市场上所有可用疗法的抵抗力的能力的阻碍(Antony和Parija,2016年)。因此,了解寄生虫获得对抗疟药的耐药性的机制对于未来替代有效治疗的发展至关重要。如今,阿耳震蛋白及其衍生物(Arts)是推荐的治疗方法,以及长期伴侣,形成基于青蒿素的联合疗法(ACTS)。artemisin抗性,主要由环阶段存活测定法(RSA)定义,经常与K13蛋白中的突变有关,而K13蛋白不调节蛋白酶体的活性(Wicht等,2020)。然而,使用蛋白酶体抑制剂(例如环氧素)会增加抗性和敏感寄生虫中的青蒿素活性(Bozdech等,2015)。在该帐户中,泛素 - 蛋白酶体途径(UPP)的不同部分的突变可能会影响阿甘辛蛋白的反应(Bridgford等,2018)。最近的研究表明,19S和20S的蛋白酶体亚基的突变敏化K13 C580Y寄生虫,这是基于RSA的更大湄公河区域中最普遍的青蒿素耐药性突变,基于RSA(Rosenthal和Ng,2021; Rossenthal和Ng,20223)。此外,在编码非素化酶UBP-1的基因中的两个突变在抗甲半氨着这甲蛋白蛋白的抗chabaudi P. chabaudi寄生虫中被鉴定出来,并且证明它们可以介导恶性疟原虫中的艺术耐药性(Cravo,2022222)。后者负责底物的识别,去泛素化,展开和易位。泛素 - 蛋白酶体系统对于真核细胞至关重要,因为它负责蛋白质的降解或回收利用,侵蚀了几个细胞过程,包括细胞周期,转录调节,细胞应激反应,信号转导,信号转导,和细胞曲折(Wang et al。,2015年)。这种蛋白质调节对于在两个宿主之间的生命周期进程中发生的疟疾寄生虫经历的快速转化至关重要,尤其是在复制率高的阶段(Krishnan和Williamson,2018年)。UPP涉及一种称为泛素化的蛋白质后修饰过程,该过程将多泛素链连接到随后由26S蛋白酶体识别的蛋白质上。如果蛋白质被蛋白质组恢复或降解,则泛素化定义的类型(Aminake等,2012; Wang等,2015)。26S蛋白酶体是一种枪管形的多亚基蛋白酶复合物,分为20S核心颗粒(CP)和19S调节粒子(RP)。20S核心通过肽基戊酰基肽水解(PGDH)(caspase样),类似胰蛋白酶样和类似chymotrypsin的活性负责蛋白水解,分别遇到了三种B-亚基(B1,B2和B5)(分别为Wang et al。,2015年)。这些催化活性的亚基分别使用N末端苏氨酸作为酸性,胰蛋白酶和疏水残基的羧基末端后的亲核试剂和裂解。这些活动站点
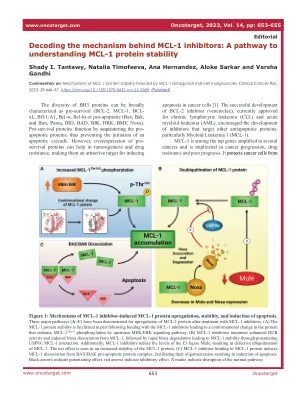
了解 MCL-1 蛋白质稳定性的途径
图 1:MCL-1 抑制剂诱导的 MCL-1 蛋白上调、稳定性和诱导细胞凋亡的机制。已证实使用 MCL-1 抑制剂治疗后,MCL-1 蛋白有三种主要上调途径 (A – C)。 (A) MCL-1 蛋白稳定性在一定程度上是在与 MCL-1 抑制剂结合后得到促进的,导致蛋白质构象变化,从而通过上游 MEK/ERK 信号通路增强 MCL-1 Thr163 磷酸化。 (B) MCL-1 抑制剂治疗增强了 DUB 活性并诱导 Noxa 与 MCL-1 解离,随后 Noxa 快速降解,通过增强 USP9x:MCL-1 相互作用实现 MCL-1 稳定性。此外,MCL-1 抑制剂降低了 E3 连接酶 Mule 的水平,导致 MCL-1 泛素化缺陷。净效应表现为 MCL-1 蛋白稳定性增加。 (C) MCL-1 抑制剂与 MCL-1 蛋白结合,诱导 MCL-1 与 BAX/BAK 促凋亡蛋白复合物分离,促进其寡聚化,从而诱导细胞凋亡。黑色箭头表示增强作用,红色箭头表示抑制作用。X 标记表示正常通路中断。

全基因组 CRISPR 筛选确定非典型 NF-kB 信号是密度依赖性增殖的调节剂
摘要上皮细胞具有维持适当细胞密度的内在机制,以实现正常组织形态形成和体内平衡。此类机制的缺陷可能导致增生和癌症发生。为了识别调节小鼠乳腺上皮细胞密度依赖性增殖的基因,我们开发了一种基于荧光泛素化细胞周期指示剂的荧光激活细胞分选检测,该检测用不同的荧光团标记细胞周期的不同阶段。利用这种强大的检测方法,我们进行了全基因组 CRISPR/Cas9 敲除筛选,筛选出在低密度下正常增殖但在高密度下继续分裂的细胞。出乎意料的是,其中一个热门结果是 Traf3,它是 NF- k B 信号的负调节剂,此前从未与密度依赖性增殖有关。我们证明 Traf3 的缺失会特异性地激活非规范的 NF- k B 信号。这反过来又触发了先天免疫反应,并通过阻止进入静止状态来驱动细胞分裂,而不依赖于已知的密度依赖性增殖机制,包括 YAP/TAZ 信号和细胞周期蛋白依赖性激酶抑制剂。

ufmylation在肿瘤发生和癌症免疫疗法中的作用
蛋白质后翻译修饰(PTMS)代表细胞调节的关键方面,在蛋白质合成mRNA后发生。这些修饰包括磷酸化,泛素化,乙酰化,甲基化,糖基化,糖基化,sumoylation和棕榈酰化,在调节蛋白质功能中起关键作用。PTM会影响蛋白质的定位,稳定性和相互作用,从而响应内部和外部刺激来策划各种细胞过程。失调与一系列疾病,例如癌症,炎症性疾病和神经退行性疾病有关。ufmylation是一种PTMS,最近因其在众多细胞过程中的调节作用而获得了突出的,包括蛋白质稳定性,对细胞应激的反应以及关键信号通路影响细胞功能。本评论强调了ufmylation在肿瘤发展和发展中的关键功能,强调了其作为治疗靶标的潜力。此外,我们讨论了ufmyration在肿瘤发生和恶性进展中的关键作用,并探索其对癌症免疫疗法的影响。本文旨在详细概述ufmylation的生物学功能,并提出靶向ufmylation如何增强癌症免疫疗法策略的有效性。

在正常和病理条件下大脑表观遗传修饰的概述
摘要:表观遗传变化是基因表达的变化,这些变化不涉及对DNA序列的改变。这些变化导致建立所谓的表观遗传密码,该代码决定了哪些基因被激活,从而策划了基因调节并在发育,健康和疾病中起着核心作用。大脑主要是由一生中不经历更新过程的细胞形成的,它很容易受到改变导致神经元死亡和神经退行性疾病的风险,主要是在较晚。在这里,我们回顾了大脑中描述的主要表观遗传修饰,并特别注意与发育异常或神经退行性疾病的发作和/或在老年发生有关的人。DNA甲基化和几种类型的组蛋白修饰(乙酰化,甲基化,磷酸化,泛素化,Sumoylation,sumoylation,lactylation和Crotonylation)是这些过程的主要参与者。他们直接或间接参与了阿尔茨海默氏病或帕金森氏病的神经变性发作。因此,本综述简要描述了这些表观遗传变化在大脑发育,成熟和衰老机制中的作用,以及动态调节或促进这些变化的一些最重要的因素,例如氧化应激,炎症和线粒体功能障碍。

CBL-B – 即将推出的免疫肿瘤学靶点
摘要 简介:E3 泛素连接酶 Cbl-b 是免疫肿瘤学中的一种新靶点,在调节 T 细胞活化和信号通路方面发挥着关键作用。通过促进关键信号蛋白的泛素化和降解,Cbl-b 调节免疫反应,维持免疫稳态并防止不必要的 T 细胞增殖。Cbl-b 作为癌症免疫治疗靶点的治疗潜力在于其对免疫抑制肿瘤微环境的贡献,目前正在努力开发小分子抑制剂。 涵盖的领域:我们回顾了 2018 年至 2024 年针对 Cbl-b 的小分子和抗体-药物偶联物。这些专利是通过公开数据库收集的,并使用手稿中描述的内部开发的化学信息学工作流程进行分析。专家意见:靶向 Cbl-b 是免疫肿瘤学的一种有前景的方法,它提供了一种新的途径来增强免疫系统对抗癌症的能力,而不仅仅是 PDL1/PD1 抑制。Cbl-b 抑制剂的开发和临床进展,正如正在进行的试验所证明的那样,标志着朝着利用这一靶点获得治疗效益迈出了重要一步。总体而言,Cbl-b 的战略抑制对改善癌症免疫治疗结果具有重大希望,预示着抗癌斗争的新时代。

紧密连接蛋白在胶质肿瘤发展和进展中的作用
紧密连接在上皮细胞和内皮细胞中形成细胞旁屏障,并调节液体、分子的扩散以及细胞在组织隔室中的渗透。紧密连接由一组整合膜蛋白组成,包括紧密连接蛋白家族、紧密连接相关 Marvel 蛋白家族、连接粘附分子家族以及锚定细胞骨架的蛋白质,例如小带闭合蛋白和扣带蛋白家族。神经递质或细胞因子等多种因素以及缺血/缺氧、炎症、肿瘤发生、磷酸化/去磷酸化、泛素化和棕榈酰化等过程调节紧密连接蛋白。紧密连接蛋白参与导致神经胶质瘤形成的肿瘤发生过程。在神经胶质瘤中,紧密连接蛋白、闭合蛋白和小带闭合蛋白-1 丰度明显失调,并且已观察到它们的错位。细胞间粘附力减弱和细胞分离是导致神经胶质瘤渗入周围组织的原因。此外,血脑屏障的旁细胞通透性(由紧密连接蛋白参与形成)会影响肿瘤周围水肿的发展,同时也会影响药物向神经胶质肿瘤的输送速度。了解脑肿瘤中的连接和旁细胞环境对于预测神经胶质肿瘤进展和化疗药物输送的可行性至关重要。这些知识也可能阐明高级别和低级别神经胶质瘤之间的差异。

肿瘤特异性 GPX4 降解增强铁死亡......
脂质过氧化依赖性铁死亡已成为一种新兴的肿瘤治疗策略。然而,目前的策略不仅选择性地诱导恶性细胞中的铁死亡,而且还同时触发免疫细胞中的铁死亡,这可能会损害抗肿瘤免疫力。在这里,我们使用 In-Cell Western 检测结合无偏药物筛选,确定化合物 N6F11 是一种铁死亡诱导剂,可触发谷胱甘肽过氧化物酶 4 (GPX4)(一种关键的铁死亡抑制剂)的降解,特别是在癌细胞中。N6F11 不会导致免疫细胞(包括树突状细胞、T 细胞、自然杀伤细胞和中性粒细胞)中的 GPX4 降解。从机制上讲,N6F11 与癌细胞中 E3 泛素连接酶三联基序 25 (TRIM25) 的 RING 结构域结合,从而触发 TRIM25 介导的 K48 连接 GPX4 泛素化,导致其蛋白酶体降解。从功能上讲,N6F11 治疗导致铁死亡癌细胞死亡,从而启动由 CD8 + T 细胞介导的 HMGB1 依赖性抗肿瘤免疫。N6F11 还增强了晚期癌症模型中针对 CD274/PD-L1 的免疫检查点阻断,包括由 KRAS 和 TP53 突变驱动的胰腺癌基因工程小鼠模型。这些发现可能建立一种安全有效的策略来增强铁死亡驱动的抗肿瘤免疫。

蛋白质Isgylation:翻译后修饰,对恶性肿瘤的影响
干扰素(IFN)刺激的基因15(ISG15)是由两个泛素样(UBL)结构域组成的15 kDa蛋白。一个铰链序列将N末端UBL结构域连接到C末端UBL结构域,该结构域具有含有赖氨酸,精氨酸和甘氨酸残基(LRLRRGG)的基序[1-4]。通过此序列,ISG15通过E1-激活酶(UBE1L)的顺序作用,E2偶联酶[泛素蛋白 - 偶联酶E2 L6(UBCH8)和E3 liig and rldl rldl rldl rldl and hh3 lig hh 3 ligh酶(ube1l)和rldl rld hhst iSG15与赖氨酸(LYS)残基上的靶蛋白共价相关。泛素蛋白连接酶5(HERC5),Ariadne RBR E3泛素蛋白连接酶1(Hhari)和包含25个(TRIM25)的三方基序[5-8]。此过程称为IsgyLation,以三个步骤发生,类似于蛋白质泛素化过程:(a)UBE1L介导了三磷酸腺苷(ATP)依赖性硫酯与ISG15的形成; (b)ISG15通过式式反应从UBE1L转移到UBCH8,形成ISG15和UBCH8之间的硫酯键; (c)从ISG15-E2酶复合物中,E3连接酶促进了ISG15向靶蛋白的LYS残基的转移和共价附着。因此,e3 ligases herc5,hhari和trim25介导底物的特异性[5-8]。蛋白质Isgylation受调节

拟南芥中脱氧酮诱导的矮人降解的新机制
在被子植物中,斯特龙酮受体是α /β水解酶dwarf14(d14),在strigolactone结合后,经历了构象变化,触发了strigolactone依赖性反应,以及strigolactones。strigolactone信号传导涉及在strigolactone结合的D14,E3-泛素li gase scf max2和转录核心代理SMXL6/7/8之间形成复合物,这些corepressors smxl6/7/8被泛素化和降级。strigolactone也破坏了D14受体的稳定性。当前模型提出D14通过SCF MAX2和蛋白酶体降解在SMXLS泛素化后发生D14降解。使用荧光和发光测定在表达与绿色荧光蛋白或荧光素酶的D14的转基因线上,我们表明,strigolactone诱导的D14降解也可能独立于SCF MAX2和/或SMXL6/7/8,通过蛋白酶体依赖性依赖性机制发生。此外,斯特龙酮水解对于触发D14或SMXL7降解不是必不可少的。还检查了突变体D14蛋白的活性,预测对斯特龙酮SIG nalling的功能是非功能的,并使用差异扫描荧光法研究了它们在体外结合Strigolactone的能力。最后,我们发现在某些条件下,D14降解的效率与SMXL7降解的效率不符。这些发现表明,与以前预期的有关D14降解的更复杂的调节机制,并提供了拟南芥信号传导动力学的新见解。