XiaoMi-AI文件搜索系统
World File Search Systembrane

ld-肽对农杆菌的适应性和极性生长至关重要
肽聚糖(PG)是一种网状结构,是细菌细胞壁的主要成分,对于维持细胞完整性和形状至关重要。大多数细菌依靠青霉素结合蛋白(PBP)进行交联,但某些物种也采用LD-转肽酶(LDTS)。与PBP不同,LDT的本质和生物学功能在很大程度上不清楚。以其极性生长而闻名的字母细菌的杂种菌序,其PG异常富含LD-Crosslinks,这表明LDT在这些细菌中可能在PG合成中起更重要的作用。在这里,我们研究了植物病原体农杆菌tumefaciens中的LDT,发现该细菌中至少有14个假定的LDT中的14种引起的LD-肽对其存活至关重要。值得注意的是,缺乏独特的7个LDT的突变体在杂种菌中广泛保守的突变体表现出降低的LD互动和PG将PG束缚到外膜β-贝尔β-桶蛋白上的链接。因此,这种突变体遭受了严重的健身损失和细胞形状的圆形,强调了这些菌粒特异性LDT在维持细胞壁完整性和促进延伸方面所起的关键作用。tn-sequering屏幕表现出了a的非冗余功能。Tumefaciens LDTS。具体而言,连字符特异性LDTs与除法和细胞周期蛋白表现出合成的遗传相互作用,而来自另一组的单个LDT。此外,我们的发现表明,缺乏所有LDT的菌株表现出独特的表型特征和遗传相互作用。总体而言,我们的工作强调了ld-rosslinking在a中的关键作用。tume-faciens细胞壁完整性和生长,并为这些交联活动的功能专业化提供了见解。

目前有几种技术用于将 RNA 分子退火到其互补的 DNA 序列。为了某些目的,RNA 和 DNA 都可以
目前有几种技术可用于将 RNA 分子与其互补的 DNA 序列退火。对于某些目的,RNA 和 DNA 都可以在溶液中,1'2 但将 DNA 固定在固体或半固体基质中,4 或附着在硝酸纤维素膜过滤器上往往更方便。5 通常在用核糖核酸酶处理以去除未杂交的 RNA 后,通过对放射性 RNA 进行闪烁计数来检测杂交体。RNA 与细胞学制剂中的 DNA 的杂交应表现出高度的空间定位,因为每种 RNA 只与其互补的序列杂交。细胞学杂交技术的一般原理并不难制定。染色体或细胞核应以尽可能逼真的方式固定;碱性蛋白质应被去除,因为它们会干扰杂交过程;5 应以不丢失细胞完整性的方式变性 DNA;杂交应使用具有极高比活度的放射性 RNA,因为在给定位点杂交的分子数很少;检测应通过氚放射自显影实现最大细胞学分辨率。本文介绍了一种适用于传统南瓜制剂的细胞学杂交技术。它以蟾蜍 Xenopus 卵母细胞中 rRNA 与染色体外 rDNA 的杂交为例。1968 年 12 月,在巴西贝洛奥里藏特举行的国际核生理学和分化研讨会上提交了该技术的初步报告。材料和方法。- 细胞学杂交技术结合了琼脂柱4 和过滤方法5 的某些特点。它应该普遍适用于任何可以作为南瓜或涂片检查的材料。制备图 1 中所示的制剂时采用以下步骤。(1)将新近变态的非洲爪蟾的卵巢在乙醇-乙酸(3:1)中固定几分钟。(2)将组织转移到显微镜载玻片上的一滴 45% 乙酸中,

MXenes作为二次电池阳极和阴极的最新进展及比较分析
MXenes 是一种寿命长达十年的陶瓷材料,于 2011 年在德雷塞尔大学首次发现 1 。它们的通式为 M n +1 X n T x , (n=1,2,3) ,其中 T 是表面终止原子,M 是早期过渡金属,X 是 C 或 N 2-4 ,MXenes 直接从其相应的 MAX 相蚀刻而成。后者是层状碳化物或氮化物结构,公式为 M n +1 AX n , (n=1,2,3) ,其中 A 是元素周期表 A 族元素,通常是第 13 或 14 族。在图 1 中,我们可以看到元素周期表中 MAX 相和 MXenes 的成分以及它们的结构。具有 OH 或 F 终端的碳化钛 Ti 3 C 2 是从钛铝 MAX 相 Ti 3 AlC 2 1 中发现的第一个 MXene。由于 Ti 3 C 2 T x MXene 仍然最具导电性 6–8,文献中对其在二次(即可充电)电池中的应用潜力进行了广泛研究。为此,人们试图通过操纵终端原子 8,9 来控制其电子和机械性能。可充电离子电池是一种基于离子插入的储能装置 10。通常,离子电池由阴极(正极)和阳极(负极)组成,并与含有离子的电解质接触。两个电极由微孔聚合物膜(隔膜)隔开,该膜阻止电子与离子一起在它们之间穿过 11。商用电池单元通常是在放电状态下生产的,而阳极和阴极电极在与大气接触时需要保持稳定 11。充电时,电极需要连接到外部电源,而电池

COVID 19感染的神经化学失调
冠状病毒之所以被命名,是因为装饰其表面的尖峰蛋白的光环[1,2]。这些S蛋白具有特定细胞受体与宿主细胞结合的特定细胞受体,然后是蛋白酶介导的S蛋白裂解,该蛋白蛋白裂解暴露了促进病毒EN尝试的融合促进域。SARS-COV-2通过其S蛋白与血管紧张素I在靶细胞上转化酶2(ACE2)Re型的血管紧张素I之间感染细胞。ACE2在肾素 - 血管紧张素系统中起关键的调节作用,该系统调节血压,盐和水平衡[3]。感染需要S蛋白质裂解,可能由宿主细胞丝氨酸蛋白酶TMPRSS2(TransMem Brane蛋白酶,丝氨酸2),尽管也可能涉及其他蛋白酶。SARS-CoV-2 belongs (Severe acute respiratory syn drome-related coronavirus 2) to the coronavirus family, which includes the pandemic MERS-CoV (Middle East respiratory syn drome coronavirus) and SARS-CoV (SARS (Severe acute respira tory syndrome)-associated coronavirus) and the lesser known but more common endemic coronaviruses HCoV-OC43 (人冠状病毒OC43),HCOV-HKU1,HCOV-229E和HCOV-NL63。特有的冠状病毒会感染上呼吸道,并频繁引起普通感冒,这反过来又与气味和味道的急性和慢性变化有关[4,5]。SARS-COV还使用ACE2作为其主要受体,在一个案例研究中,SARS-COV感染急性病毒介导的气味变化的主要机制包括由于粘膜肿胀而导致通气损失引起的导电缺陷,粘液产生增加,粘液组成的变化,粘液组成的变化以及嗅觉信号的次要变化以及局部释放的诸如colied coilsied coil synemiss andery机制的局部释放引起的嗅觉信号的变化,导致多种机制的释放,导致多种机制释放,导致其造成的流行。嗅觉缺陷倾向于使用类似于其他与冷相关的Symp Toms(如鼻充血)和HCOV-NL63(HCOV-NL63)相似的时间过程,而特有的冠状病毒不将ACE2用作其主要的细胞受体[6],这是一种可能基本的分子诊断,可能是致病物理学中关键差异的基础。

线粒体丙酮酸载体抑制剂的最新进展
在真核细胞中,线粒体是内共生器官,与各种细胞过程有关,包括能量消耗,生物合成,信号转移和程序性细胞死亡。1显着,它们是创建三磷酸腺苷(ATP)的主要位置,腺苷三磷酸腺苷(ATP),包括所有生物的通用自由能载体,包括所有五个呼吸链络合物和所有三羧酸周期(TCA)酶。在细胞质和线粒体基质之间的代谢物交换对于执行这些代谢过程是必要的,这些代谢过程仅限于线粒体腔室并保留内部内稳态。电压依赖性阴离子通道允许微小的分子穿过外部线膜。然而,线粒体内膜(IMM)对分子和离子高度渗透,必须依靠特定的转运蛋白和通道来连接细胞质和线粒体的代谢。线粒体载体家族成员执行大部分运输步骤。2其他转运蛋白家族包括线粒体丙酮酸载体(MPC)。3 MPC是一种蛋白质复合物,存在于线粒体内膜中,并负责将丙酮酸从线粒体转运到线粒体基质中,其中丙酮酸转化为乙酰基氧乙烯酶A(乙酰辅酶A)。ace-tyl-coa进入TCA循环,并在其中进一步氧化。另外,线粒体中的丙酮酸也可以通过吡二酸酯羧化酶的羧化来参与糖异生,以产生草乙酸以补充TCA循环。7如上所述,除了被运输到线虫外,丙酮酸还可以通过细胞质中的乳酸脱氢酶(LDH)还原为乳酸。MPC是在1970年代4提出的,最初被称为BRP44L(脑蛋白44样)和BRP44(脑蛋白44)。它在2003年被鉴定在酵母中,并在2012年进一步鉴定在哺乳动物中。3,5,6 MPC是一个相对较小的杂物,由两个亚基组成,分别由12和14 kDa组成,分别为12和14 kDa。
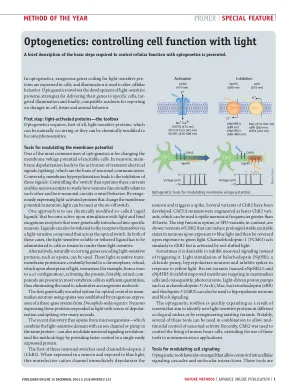
光遗传学:用光控制细胞功能
调节膜电位的工具 光遗传学最常见的用途之一是改变可兴奋细胞的膜电位。在神经元中,膜去极化会导致瞬态电信号(脉冲)的激活,这是神经元通讯的基础。相反,膜超极化会导致这些信号的抑制。控制操作这些电流的“开关”使神经科学家能够研究神经元在功能上如何相互关联以及神经元回路如何控制行为。通过外源表达改变神经元膜电位的光激活蛋白,光可以用作开关。一种方法是使用化学修饰的所谓“笼状配体”,这些配体在光刺激下变得活跃并与通过基因引入特定神经元的外源性受体结合。配体也可以通过充当光开关的光敏化合物与受体本身相连。在这两种情况下,都必须将光敏的可溶性或束缚配体注入细胞或组织,使它们对光敏感。或者,可以使用编码光敏蛋白(如视蛋白)的天然基因。这些光敏跨膜蛋白与发色团视网膜共价结合,视网膜吸收光后发生异构化(例如,从反式变为顺式构型),从而激活蛋白质。值得注意的是,视网膜化合物在大多数脊椎动物细胞中含量充足,因此无需注入外源分子。第一个利用视蛋白进行哺乳动物神经元光学控制的遗传编码系统是通过外源表达果蝇的三基因系统建立的。表达这些蛋白质的神经元对光的反应是数秒内的去极化和尖峰波。最近发现,微生物中的视蛋白(将光敏域与同一蛋白质中的离子通道或泵相结合)也可以调节神经元信号,通过在单个易于表达的蛋白质中提供更快的控制,彻底改变了该方法。这些神经元开关中的第一个使用了通道视紫红质-2 (ChR2)。当在神经元中表达并暴露于蓝光时,这种非选择性阳离子通道会立即使神经元去极化

单元格使用分子工作记忆在...
b,EGFR-PTP相互作用网络的方案。配体EGFR(E P)与PTPRG(P RG)和PTPN2(P n 2)相互作用。配体EGFR(E -E P)促进E p的自催化。因果链接 - 纯黑线;弯曲的箭头线 - 扩散,PM-质膜,ER-内质网。另见图1-图1B。C,在细胞极化过程中信号诱导的形状变化。箭头:局部边缘速度方向。Zoom:细胞的粘弹性模型 - 弹性和粘性元件的平行连接。P总计:总压力; V:当地的内存速度; L:粘弹性状态。粗字母:向量。细胞膜轮廓:[0,2π]。d,顶部:空间EGF分布的计算机演变。底部:EP的Kymograph for Handomation在(b)中网络的反应扩散模拟中的临界性。三角形 - 梯度硬化。e,用(c)中的模型获得的颜色编码E P的相应示例性细胞形状。f,顶部:颞pro纤维e p(黑色)和e -e p(灰色)。绿色阴影区域:EGF梯度存在。底部:具有表示捕获状态空间区域(彩色)和相应时间尺度的系统的状态空间轨迹。另请参见图1-视频1。厚/细线:信号前置/缺失。g,在e示例中的硅细胞形态变化中的定量变化。三角形 - 梯度持续时间。h,左:与G中相同,只有从同一方向用两个连续的动态梯度(三角形)刺激时。第二梯度在第一个的内存阶段。另见图1-图1d。右:第二个梯度(橙三角形)的方向相反。另见图1-图补充1E。虚线:g。平均值±S.D.显示了n = 3。参数:方法。在(D-H),绿色(橙)/红线:刺激存在/不存在。

环境心理学杂志
电抽搐治疗(ECT)是晚期抑郁症(LLD)的有效治疗方法,但其确切机制尚未完全了解。神经可塑性假设表明,ECT通过诱发导致神经营养作用的癫痫发作而起作用[1]。支持神经塑性假说,临床前研究表明,电性抽搐刺激(ECS)增强了神经营养因素,促进神经发生,增强突触发生,轴突发芽,树突状生长,棘突密度,以及在Hippo Campus中的突触循环(Alter)[2]。这些变化,特别是在海马和额叶前皮层中,被认为是ECT的情绪改善效果的基础[3]。然而,这些临床前发现向人类ECT治疗的翻译尚待验证。临床研究为神经可塑性假设提供了有限的直接证据,主要依赖于间接措施,例如MRI扫描来显示ECT对增加灰质体积(GMV)的影响[4]。尚不清楚这些结构变化背后的确切分子机制及其与ECT有效性的关系。此外,在ECT之后在人类中观察到的GMV的增加尚无共识,直接与所属于ECS的动物模型中看到的神经塑性变化相关。基于LeviterAcetam的新型放射性体已推动了突触密度的体内PET成像,这些放射性体的靶向具有高亲和力的靶向突触囊泡蛋白2A(SV2A)[5]。sv2a是一种关键的突触前囊泡内膜蛋白,几乎在几乎所有大脑突触中都发现[6]。a由于其SV2A亲和力和分布体积,11 C-UCB-J示踪剂是评估体内突触密度的可靠标记[5]。使用该示踪剂现在可供人类使用,研究人员可以直接观察到接受ECT的患者突触密度的变化。这项研究旨在通过利用体内突触密度成像来验证EC的临床前突触发生结果,以检测晚期抑郁症患者(LLD)患者的ECT诱导的突触发生。主要目标是评估通过T1加权MRI检测到的GMV的增加,与在急性ECT之后通过11 c-UCB-J PET测量的突触den sity的同时增加了突触den性。

蜂胶化合物作为潜在的MMP1和MMP2抑制剂的活性在伤口愈合应用中的硅研究中
基质金属蛋白酶(MMP)是一种内肽,参与了伤口愈合的所有阶段。在炎症期间,MMP去除所有受损的蛋白质和临时ECM。在增殖阶段,MMPS降解毛细血管基底膜以促进血管生成和细胞迁移。同时,在组织重塑中,MMP活性降低并诱导重塑生长因子的释放。在这种情况下,金属蛋白酶(TIMP)的组织抑制剂通过与特定位点结合并防止ECM分解过度在平衡MMP活动中起关键作用(Ayuk等,2016; Kandhwal等人,2022年)。但是,在某些条件下,MMP和TIMP的失衡导致康复过程不佳。MMP1是一种显着的colla-基因,在血管起源期间在伤口部位表达。它在伤口愈合中重新排斥和迁移后,它作为真皮的重要组成部分降解。MMP1活动仅在关闭伤口之前有效,然后在重塑阶段自动关闭。然而,高水平的MMP-1与慢性伤口相关,并导致愈合时间延长(Muller等,2008)。例如,糖尿病足溃疡患者的MMP1大幅增加。MMP1/TIMP1的比率用作糖尿病足溃疡伤口愈合的预测因子。虽然比TIMP1的比率更高,但治疗效果越糟。先前的研究发现,慢性伤口渗出液的MMP水平高于急性伤口渗出液(Lobmann等,2002)。MMP12,一种金属弹性酶,在伤口愈合中也起着重要作用。它会破坏细胞外基质弹性蛋白,并使负责发炎和肉芽肿发育的免疫细胞浸润。MMP12对1型糖尿病患者的轻度炎症产生了影响。MMP12与1型糖尿病患者的轻度炎症有关,并在1型糖尿病患者的肠道损伤中与腹腔疾病呈正相关(Bister等,2005)。

PSMA 指导的涎腺肿瘤成像和治疗:一项单中心回顾性研究
我们分析了前列腺特异性膜抗原 (PSMA) PET/CT 的诊断性能以及 177 Lu-PSMA-617 放射性配体治疗 (RLT) 在唾液腺恶性肿瘤 (SGM) 中的剂量、疗效和安全性。方法:我们从数据库中确定了 28 名接受 PSMA PET/CT 检查的 SGM 患者。CT 和 PSMA PET/CT 图像由 3 名匿名阅读者在联合阅读会话中分别评估。病理学发现分为 6 个 TNM 区域,病变范围分为无疾病(n=1,4%)、单灶性(n=2,7%)、寡转移性(n=9,32%)、多灶性(n=3,11%)或播散性(n=13,47%)。对于每个区域,测量摄取量最高的病变的 SUV max,并使用 PROMISE 标准对每个患者进行视觉 PSMA 表达评分。使用 Student t 检验测试 PSMA 表达与临床和组织病理学标志物之间的关联。五名患者接受了治疗内剂量的 PSMA RLT。使用 RECIST 1.1 评估疗效,并根据《不良事件常用术语标准》5.0 版对不良事件进行分级。结果:与 CT 相比,PSMA PET/CT 在 28 名患者中的 11 名(39%)显示了额外的转移性病变,导致 3 名(11%)和 6 名(21%)患者的 TNM 分期和病变范围上调。PSMA PET/CT 分别在 1 名(4%)、4 名(14%)、2 名(7%)和 4 名(14%)患者中检测到 CT 隐匿的局部肿瘤、区域淋巴结、非区域淋巴结和骨转移;在其他预定区域未检测到额外的病变。6 名患者(25%)的 PSMA 表达水平高于肝脏。男性患者的 SUV max 明显高于女性患者(15.8 vs. 8.5,P = 0.007),且骨骼病变的 SUV max 明显高于肺部病变(14.2 vs. 6.4,P = 0.006)。5 名患者中有 3 名在 1 个周期后停止 PSMA RLT,原因是肿瘤剂量不足。未发生 4 级或更高级别的不良事件。结论:在 SGM 中,与 CT 相比,PSMA PET/CT 显示出更高的检出率,并导致约三分之一的患者分期上升。男性和存在骨转移与 PSMA 表达明显较高有关。PSMA RLT 耐受性良好,但由于肿瘤剂量不足,大多数患者未接受超过 1 个周期的治疗。