XiaoMi-AI文件搜索系统
World File Search Systemchromosome

研究文章 NUT 中线癌:一种以染色体重排为特征的罕见实体肿瘤
目的 . 罕见低分化NUT中线癌(NMC)是一种高度恶性肿瘤。但由于NMC罕见,关于其临床、影像学和病理特征的报道仍然很少。方法 . 本研究以3例位于腮腺、肺和气管的NMC患者为例,总结NMC的临床病理特征。所有病例均通过双色FISH检测后检测NUT抗体核反应阳性进行诊断,结果均为阳性,提示NUT基因15q14发生染色体重排。结果 . 这3例患者均接受了手术治疗和放化疗等常规治疗。鉴于常规强化治疗效果不佳,建议使用两种新型疗法,即组蛋白去乙酰化酶抑制剂 (HDACi) 和溴结构域抑制剂 (BETi),因为两者都可以抑制肿瘤细胞的生长,这些靶向疗法可能会延长患者的生存时间。结论。NMC 是一种容易误诊且预后不良的癌症;因此,提高临床医生的认识对于提高诊断准确性至关重要,选择有效的治疗方法是改善预后的主要方法。

性染色体的离职额在维持柳树后的障碍中起着重要作用
salix属(柳树)中的几乎所有物种都是富裕的,柳树具有可变的性别确定系统,这种变量在维持物种屏障中的作用相对未经测试。我们首先分析了两个物种的性别确定系统(SDS),即Salix Cardiophylla和Salix Interior,它们在Salix系统发育中的位置使它们对于理解亲属中发现的性染色体更换的立场至关重要,并且从其亲属中发现了性染色体,这将系统从雄性(XX/XY)转换为女性(ZW/ZZ/ZZ)heterogamemety。我们表明,这两个物种均具有男性异质体,在15号染色体上具有性别连接区域(SLR)(称为15倍系统)。SLR分别占整个参考染色体的21.3%和22.8%。通过构造系统发育树,我们确定了所有具有已知SDS的物种的系统发育位置。祖先SDS角色状态的重建表明,15倍系统可能是柳树中的祖先状态。的15倍至15zW和15倍至7倍的失误可能导致了Salix的早期形态,并引起了Vetrix和Salix进化枝的主要组。最后,我们根据常染色体和SLR分别测试了系统发育树中物种之间的渗透率。在15倍,15zW和7倍的物种中观察到,显示出降低的渗入,尤其是15zW和7倍物种之间的基因流动较少。 我们认为,尽管柳树物种形成中的SDS失误可能不会产生完全的生殖屏障,但SLR的演变在防止渗入和维持物种边界方面起着重要作用。,显示出降低的渗入,尤其是15zW和7倍物种之间的基因流动较少。 我们认为,尽管柳树物种形成中的SDS失误可能不会产生完全的生殖屏障,但SLR的演变在防止渗入和维持物种边界方面起着重要作用。显示出降低的渗入,尤其是15zW和7倍物种之间的基因流动较少。我们认为,尽管柳树物种形成中的SDS失误可能不会产生完全的生殖屏障,但SLR的演变在防止渗入和维持物种边界方面起着重要作用。

X染色体超表达和雄性组织中灭活的动力学
。cc-by 4.0国际许可证(未经同行评审证明)获得的是作者/资助者,他已授予Biorxiv授予Biorxiv的许可,以永久显示预印本。这是该版本的版权所有,该版本发布于2024年7月3日。 https://doi.org/10.1101/2024.07.01.601468 doi:Biorxiv Preprint

大鼠染色体4的表观遗传变化有助于酒精饮酒和-nonpreferring大鼠的不同饮酒行为
*通讯作者。印第安纳大学医学院印第安纳大学医学院印第安纳大学医学院,印第安纳波利斯,印第安纳州,46202,美国。 电话。 :+1 317 2747813。传真:+1 317 274 2695.,tliang@iu.edu(T。liang)。 作者信用声明John Paul Spence:概念化;方法论;写作 - 原始制图,审查和编辑。 dongbing lai:软件;正式分析;写作 - 审查和编辑。 Jill L. Reiter:概念化;写作 - 审查和编辑。 SHA CAO:正式分析。 Richard L. Bell:方法论;正式分析;资源;写作 - 审查和编辑。 肯特·E·威廉姆斯:正式分析;写作 - 审查和编辑;可视化。 领带:概念化;监督;资源;写作 - 审查和编辑;可视化。印第安纳大学医学院印第安纳大学医学院印第安纳大学医学院,印第安纳波利斯,印第安纳州,46202,美国。电话。:+1 317 2747813。传真:+1 317 274 2695.,tliang@iu.edu(T。liang)。作者信用声明John Paul Spence:概念化;方法论;写作 - 原始制图,审查和编辑。dongbing lai:软件;正式分析;写作 - 审查和编辑。Jill L. Reiter:概念化;写作 - 审查和编辑。SHA CAO:正式分析。Richard L. Bell:方法论;正式分析;资源;写作 - 审查和编辑。 肯特·E·威廉姆斯:正式分析;写作 - 审查和编辑;可视化。 领带:概念化;监督;资源;写作 - 审查和编辑;可视化。Richard L. Bell:方法论;正式分析;资源;写作 - 审查和编辑。肯特·E·威廉姆斯:正式分析;写作 - 审查和编辑;可视化。领带:概念化;监督;资源;写作 - 审查和编辑;可视化。
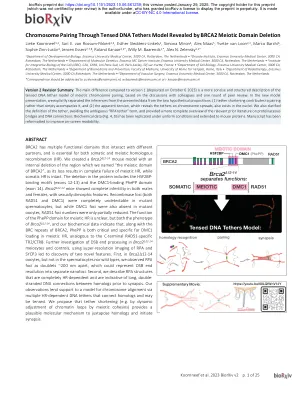
BRCA2 减数分裂域缺失揭示的通过张紧 DNA 束缚模型进行染色体配对
1 荷兰鹿特丹伊拉斯姆斯大学医学中心发育生物学系,邮编 3000 CA • 2 荷兰鹿特丹伊拉斯姆斯大学医学中心 Oncode 研究所,邮编 3000 CA • 3 荷兰鹿特丹伊拉斯姆斯大学医学中心伊拉斯姆斯 MC 癌症研究所分子遗传学系,邮编 3000 CA • 4 细胞整合生物学研究所 (I2BC)、CEA、CNRS、Uni Paris-Sud、Uni Paris-Saclay、法国吉夫河畔伊维特 • 5 荷兰鹿特丹伊拉斯姆斯大学医学中心细胞生物学系,邮编 3000 CA • 6 意大利罗马第二大学医学院生物医学与预防系 • 7 荷兰鹿特丹伊拉斯姆斯大学医学中心放射治疗系,邮编 3000 CA • 8 荷兰鹿特丹伊拉斯姆斯大学医学中心血管外科系, 3000 CA,鹿特丹,荷兰

现代人类的大脑发育中期比尼安德特人更长,染色体分离错误更少
自从现代人的祖先与尼安德特人的祖先分离以来,大约 100 种氨基酸替换传给了几乎所有现代人。这些变化的生物学意义在很大程度上是未知的。在这里,我们研究了三种蛋白质中的所有六种氨基酸替换,这三种蛋白质已知在动粒功能和染色体分离中起关键作用,并在发育中的新皮质的干细胞中高度表达。当我们将这些现代人类特异性替换引入小鼠时,其中两种蛋白质 KIF18a 和 KNL1 中的三种替换会导致中期延长,并减少发育中皮质顶端祖细胞的染色体分离错误。相反,祖先替换会导致人类脑器官中的中期长度更短和染色体分离错误更多,与我们在黑猩猩脑器官中发现的情况类似。这些结果表明,现代人类与尼安德特人分化后,大脑皮层发育过程中染色体分离的保真度得到了提高。

校正:六价铬的致癌机制:从DNA断裂到染色体不稳定性和肿瘤转化
1位环境和遗传毒理学实验室,路易斯维尔大学药理学与毒理学系,500 S Preston ST,RM 1422,RM 1422,美国肯塔基州路易斯维尔1位环境和遗传毒理学实验室,路易斯维尔大学药理学与毒理学系,500 S Preston ST,RM 1422,RM 1422,美国肯塔基州路易斯维尔

复制应力产生独特的DNA拷贝数改变和染色体量表损失
抽象背景:癌症染色体不稳定性的主要驱动力是复制应力,DNA复制的减慢或失速。尚不清楚如何连接复制应力和基因组不稳定性。蚜虫蛋白诱导的复制应力会在常见的脆弱部位诱导分裂,但是易于脆弱的确切原因,并且没有充分探索复制应力的急性基因组后果。结果:我们表征单个二倍体非转化细胞中的DNA拷贝数改变(CNA),这是由一个细胞周期在蚜虫或羟基脲存在下引起的。产生了多种类型的CNA,与不同的基因组区域和特征相关,观察到的拷贝数景观在蚜虫蛋白和羟基脲诱导的复制应力之间是不同的。将CNA与基因表达和单细胞复制时间分析的耦合细胞类型分析指向蚜虫中最复发的染色体尺度CNA的致病性大基因。这些在RPE1上皮细胞中的7号染色体上聚集在染色体上,但染色体在BJ成纤维细胞中。染色体臂水平CNA还会产生含有这些染色体的染色质和微核。结论:由复制应力驱动的染色体不稳定性通过局灶性CNA和染色体臂尺度的变化发生,后者仅限于很小的子集染色体区域,潜在地倾斜了癌症基因组的进化。复制应力的不同诱导者导致独特的CNA景观,从而提供了机会,从而得出了特定复制应力机械的拷贝数签名。单细胞CNA分析揭示了复制应力对基因组的影响,从而提供了对癌症中染色体不稳定性的分子机制的见解。

标题:染色体 10q24.32 变异与不同人群的脑动脉 1 直径相关:全基因组关联研究 2
作者:Minghua Liu 1、Farid Khasiyev 2、Sanjeev Law 1,3,4、Antonio Spagnolo-Allende 1、3 Danurys L Sanchez 1,3,4、Howard Andrews 5、Qiong Yang 6、Alexa Beiser 6、Ye Qiao 7、Emy A 4 Thomas 8、Jose Rafael Romero 9、Ta tjana Rundek 10,11,12、Adam Brickman 1,3,4、Jennifer J Manly 1,3,4 5、Mitchell SV Elkind 1,13、Sudha Seshadri 9,14、Christopher Chen 15、Ralph L Sacco 10,11,12、Saima 6 Hilal 15、Bruce A Wasserman 7,16、Giuseppe 1,3,4、Myriam Fornage 8,17; 7 8 隶属关系: 9 1 哥伦比亚大学瓦格洛斯内科与外科医学院神经病学系,10 美国纽约州纽约市 11 2 圣路易斯大学医学院神经病学系。密苏里州路易斯,12 3 TAUB研究疾病研究所和衰老大脑,Vagelos学院,13个医师和外科医生,哥伦比亚大学,纽约,纽约,纽约,14 4 4 4 The Sergievsky Center,Vagelos医师和外科学院,哥伦比亚大学,15年,纽约大学,纽约大学,纽约,美国16 5 BISTATIS IS CUPPLY SHILLECH,MAIL SHICOL,MAIL SHILEBY,USY SEPRICY,MA SERVELY,MA NOWSONN,17 NY SONTON,BOST,NY ny约翰·霍普金斯大学医学院,美国马里兰州20 8布朗基金会分子医学研究所,MC政府医学院,美国德克萨斯州霍斯顿市德克萨斯大学卫生科学中心21 2 22 9美国马萨诸塞州波士顿医学院神经病学部美国佛罗里达州27号Iami Miller医学院28 13 13哥伦比亚大学哥伦比亚大学,美国新纽约州30 14 Glenn Biggs阿尔茨海默氏症和神经退行性疾病研究所,德克萨斯大学健康科学中心,美国德克萨斯州圣安东尼奥市31号

8p染色体工程揭示了肝癌中患者特异性合成致死性靶向的转移潜力增加
大规模的染色体畸变在人类癌症中普遍存在,但其功能仍然很差。我们使用CRISPR-CAS9基因组编辑建立了染色体工程的肝细胞癌细胞系。A 33 - Mega - 8p染色体(CHR8P)上的碱基对区域被删除,模仿了一个经常观察到的染色体缺失。使用此等源性模型系统,我们描述了CHR8P损失的功能序列及其对转移行为和患者生存的影响。我们发现,CHR8P的转移基因协同起作用,以诱导CHR8P骨骼肿瘤的侵略性和侵入性表型的特征。全基因组CRISPR-CAS9在等源性CHR8P删除细胞中的可行性筛选是一种强大的工具,可以找到先前未识别的合成致死靶标和伴随患者特异性染色体改变的脆弱性。使用此目标识别策略,我们表明CHR8P的de依将肿瘤细胞敏感到靶向反应性氧消毒酶Nudix水解酶17。因此,综合工程允许鉴定新型的合成致死性,特异性杂志的杂合性丧失。