XiaoMi-AI文件搜索系统
World File Search Systemenables
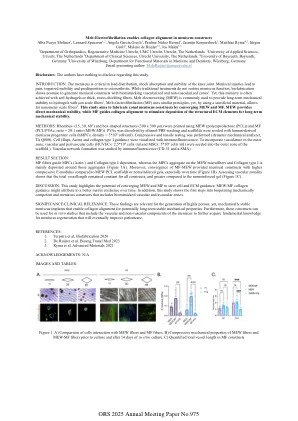
熔融电颤技术使半月板结构中的胶原蛋白排列整齐
披露:作者对于本研究没有任何需要披露的信息。简介:半月板对于膝关节的负荷分布、减震和稳定性至关重要。半月板损伤会导致疼痛、活动受限和易患骨关节炎。虽然传统治疗方法不能恢复半月板功能,但生物制造有望生成具有仿生血管化和非血管化区域的半月板结构 1 。然而,这种模拟通常是通过软水凝胶或厚的应力屏蔽纤维实现的。熔融电写 (MEW) 通常用于为具有 µ m 级纤维的水凝胶提供长期机械稳定性 2 。熔融电纤颤 (MF) 使用类似原理,但通过使用牺牲材料,可以实现纳米级纤维 3 。本研究旨在通过融合 MEW 和 MF 来制造区域性半月板结构。 MEW 提供直接的机械稳定性,而 MF 引导胶原蛋白排列以刺激结构 ECM 元素的沉积,从而实现长期的机械稳定性。方法:使用 MEW(聚己内酯 (PCL))和 MF(PCL/PVAc,比例 = 20:1(MEW:MF))打印菱形(15、30、60 °)和盒子状结构(300 x 300 µm)。通过乙醇/PBS 洗涤溶解 PVAc,并在支架上接种人源半月板祖细胞(hMPC,密度 = 5*10 6 细胞/毫升)。进行压缩和拉伸测试(动态机械分析仪,TA Q800)。用免疫荧光可视化细胞(Dapi、肌动蛋白)和 I 型胶原蛋白引导。为了将脉管系统纳入外部区域,将血管和血管周围细胞(HUVEC:2.5*10 6 细胞/ml 和 MSC:5*10 6 细胞/ml)接种到支架的外部区域。)通过免疫荧光(CD-31 和 a-SMA)研究血管网络的形成。结果部分:MF 纤维引导 MPC(肌动蛋白 +)和 I 型胶原蛋白沉积,而 MPC 聚集在 MEW 微纤维上,I 型胶原蛋白主要沉积在这些聚集体周围(图 1A)。此外,与 MEW PCL 支架或非增强凝胶相比,MF-MEW 的汇聚为半月板结构提供了更高的压缩 E 模量,尤其是随着时间的推移(图 1B)。评估血管分区显示所有结构的总血管长度保持不变,并且与非增强凝胶相比更大(图 1C)。讨论:本研究强调了 MEW 和 MF 融合以引导细胞和 ECM 引导的潜力。MEW/MF 胶原引导可能归因于随着时间的推移更好的基质弹性。此外,本研究展示了生物打印机械能力和半月板构造的第一步,其中包括仿生血管和无血管区。意义/临床意义:这些发现与生成高度多孔但机械稳定的半月板植入物有关,这些植入物可实现胶原对齐,从而实现潜在的长期稳定机械性能。此外,这些结构可用于包括半月板血管和非血管成分的体外研究,以进一步获得半月板再生的基础知识,最终改善患者护理。参考文献:

高质量基因组组装使得能够预测等位基因...
1 北京林业大学生物科学与技术学院, 国家林木育种与生态修复工程研究中心, 林木分子设计育种北京市高精尖创新中心, 林木育种国家工程实验室, 林木园林植物遗传育种教育部重点实验室, 北京 100083 2 山东省农业科学院作物种质资源研究所, 作物遗传改良与生理生态重点实验室, 济南 250100 3 广东省农业科学院水稻研究所, 农业农村部南方优质水稻遗传育种重点实验室 (省部共建) , 广东省水稻育种新技术重点实验室, 广州 510640 4 宁夏大学农学院, 银川 750021 5 云南省水稻遗传改良重点实验室中国科学院昆明植物研究所东亚植物多样性与生物地理学重点实验室极小种群植物综合保护重点实验室,云南昆明 650201 6 山东农业大学林学院,山东泰安 271000 7 于默奥大学生态与环境科学系于默奥植物科学中心,瑞典于默奥 SE-901 87 8 不列颠哥伦比亚大学林业与保护科学系,加拿大不列颠哥伦比亚省温哥华,V6T 1Z4 9 图能森林遗传研究所,德国格罗斯汉斯多夫 22927 10 根特大学植物生物技术和生物信息学系,比利时根特 9052 11 VIB 植物系统生物学中心,比利时根特 9052 12 微生物生态学和基因组学中心,比勒陀利亚大学生物化学、遗传学和微生物学系,比勒陀利亚 0028,南非 13 南京农业大学园艺学院,高等交叉研究院,南京 210095,中国 14 于默奥植物科学中心,植物生理学系,于默奥大学,SE-901 87 于默奥,瑞典 15 森林与森林科学系,Faculté de林业,地理与地理,拉瓦尔大学,魁北克,QC G1V 0A6,加拿大

布洛赫方程使物理信息神经网络成为可能......
摘要 — 磁共振成像 (MRI) 是临床诊断中一种重要的非侵入性成像方法。除了常见的图像结构之外,参数成像还可以提供内在的组织特性,因此可用于定量评估。新兴的深度学习方法提供了快速准确的参数估计,但仍然缺乏网络解释和足够的训练数据。即使有大量的训练数据,训练数据和目标数据之间的不匹配也可能导致错误。在这里,我们提出了一种仅依赖于目标扫描数据而不需要预定义训练数据库的方法。我们提供了一个概念验证,将 MRI 的物理规则 Bloch 方程嵌入到物理信息神经网络 (PINN) 的损失中。PINN 能够学习 Bloch 方程,估计 T2 参数,并生成一系列物理合成数据。在幻像和心脏成像上进行了实验结果,以证明其在定量 MRI 中的潜力。

修复CRISPR引导的RNA断裂启用站点 -
。STHCSM的RNA裂解在每个切割位点生成2',3'> P和5'-OH,我们假设RTCB的连接酶活性参与了图中确定的RNA修复。1(中间)。b)用抗RTCB或抗ACTB(加载对照)抗体的蛋白质印迹,用(+)或没有( - )RTCB耗竭的293T细胞的裂解物进行抗体。参见图中的未编写图像。S2。c)PARK7成绩单针对5

我们创造空间,让非凡之事得以发生
我们的负责任 SEGRO 框架侧重于三个长期优先事项,我们相信我们可以为这些优先事项做出最大的商业、环境和社会贡献。对于每个领域,我们都设定了具有挑战性的初始目标,并每年报告这些目标,并制定了实现这些目标所需的行动。我们将根据需要设定额外的、更具体的支持目标,我们希望我们的行动和方法能够随着时间的推移而发展,以反映我们的成就、技术变革以及我们的利益相关者和更广泛社会的优先事项。
crispr可为绿色未来
群集定期间隔短的短文重复序列(CRISPR)/ CRISPR相关蛋白9(CAS9)系统已成为过去十年来许多动植物和动物物种中靶向基因组编辑的最重要工具。CRISPR/CAS9技术还引发了关键谷物作物中基因组编辑的应用和技术进步的范围,包括大米,小麦,玉米和大麦。在这里,我们回顾了CRISPR/ CAS9的先进用途以及在谷物作物的基因组编辑中的衍生系统,以增强各种农艺重要特征。我们还重点介绍了提供的新技术进步,用于提供预组装的cas9-grna核糖核蛋白(RNP)编辑系统,多重编辑,功能获得的策略,使用人工智能(AI)工具(AI)工具以及将CRISCR与新型快速繁殖(SB)结合在一起(SB)和Vernalsization and Nalnalization and Nalnalization Caltergies。

上游开放阅读框的基因组编辑启用...
通过上游开放式阅读框(UORF)的翻译调节已成为控制从下游初级ORF(PORFS)1-5合成的蛋白质量的一般机制。我们发现,植物中内源性UORF的基因组编辑能够调节来自四个porfs的mRNA的翻译,这些porfs涉及发育或抗氧化生物合成。针对具有UORF启动密码子的区域的单个指定RNA可以产生多个突变。UORF编辑后,我们观察到四个porfs中的mRNA翻译数量不同。值得注意的是,编辑了LSGGP2的UORF,该UORF在生菜中编码了维生素C生物合成的关键酶,不仅增加了氧化应激耐受性,而且增加了抗坏血酸含量的含量约150%。这些数据表明,编辑植物UORFS提供了一种可推广,有效的方法来操纵mRNA的翻译,可用于剖析生物学机制并改善农作物。

通过Fullerene Blends优化接口启用 - ...
Interface Optimization via Fullerene Blends Enables Open-Circuit Voltages of 1.35 V in CH 3 NH 3 Pb(I 0.8 Br 0.2 ) 3 Solar Cells Zhifa Liu # , Johanna Siekmann # § , Benjamin Klingebiel, Uwe Rau, and Thomas Kirchartz* § Dr. Z. Liu, J. Siekmann, Dr. B. Klingebiel, Prof. U. Rau,T。KirchartzIek5-Photovoltaik教授,ForschungszentrumJülich,52425Jülich,德国#作者贡献Z.L.和J.S.同等贡献。*教授。 T. Kirchartz工程学院和Cenide,Carl-Benz-STR的Duisburg-Essen大学。199,47057德国杜伊斯堡§相对作者电子邮件:t.kirchartz@fz-juelich.de电子邮件:

工程CRISPR/CAS12A启用快速SARS-COV-2检测
致谢:我们感谢Jain Lab的成员提供有用的建议和讨论。我们感谢佛罗里达大学,UF健康癌症中心和临床和转化科学研究所存储库的支持。我们还要感谢David Ostrov博士和Cuong Nguyen博士的支持,并获得了患者样本并进行讨论。 最后,我们感谢惠特尼·斯托佩尔(Whitney Stoppel)博士和她在化学工程系的实验室和佛罗里达大学森林基因组学集团的Chrisopher Dervinis博士使用冻干剂。 通过BEI资源,NIAID,NIH:中东呼吸综合征冠状病毒(MERS-COV),EMC/2012,NR-45843获得以下试剂。来自Urbani,Urbani,NR-52346的定量PCR(QPCR)控制RNA;来自人类冠状病毒(HCOV)的基因组RNA,NL63,NR-44105;来自SARS相关冠状病毒2的基因组RNA,孤立的香港/VM20001061/2020,NR-52388。 以下试剂是由疾病控制和预防中心沉积的,并通过BEI资源,NIAID,NIH:与SARS相关冠状病毒2,分离的USA-WA1/ 2020,NR-52285的BEI资源:基因组RNA。来自热灭活SARS相关的冠状病毒2,分离株USA-WA1/2020,NR 52347的定量PCR(QPCR)对照RNA;与SARS相关的冠状病毒2,分离的USA-WA1/2020,热灭活,NR-52286。 通过Bei Resources,NIAID,NIH:与SARS相关的冠状病毒2,Inalaly Italy-Inmi1,NR-52498分离出Maria R. Capobianchi博士的基因组RNA。我们还要感谢David Ostrov博士和Cuong Nguyen博士的支持,并获得了患者样本并进行讨论。最后,我们感谢惠特尼·斯托佩尔(Whitney Stoppel)博士和她在化学工程系的实验室和佛罗里达大学森林基因组学集团的Chrisopher Dervinis博士使用冻干剂。通过BEI资源,NIAID,NIH:中东呼吸综合征冠状病毒(MERS-COV),EMC/2012,NR-45843获得以下试剂。来自Urbani,Urbani,NR-52346的定量PCR(QPCR)控制RNA;来自人类冠状病毒(HCOV)的基因组RNA,NL63,NR-44105;来自SARS相关冠状病毒2的基因组RNA,孤立的香港/VM20001061/2020,NR-52388。以下试剂是由疾病控制和预防中心沉积的,并通过BEI资源,NIAID,NIH:与SARS相关冠状病毒2,分离的USA-WA1/ 2020,NR-52285的BEI资源:基因组RNA。来自热灭活SARS相关的冠状病毒2,分离株USA-WA1/2020,NR 52347的定量PCR(QPCR)对照RNA;与SARS相关的冠状病毒2,分离的USA-WA1/2020,热灭活,NR-52286。通过Bei Resources,NIAID,NIH:与SARS相关的冠状病毒2,Inalaly Italy-Inmi1,NR-52498分离出Maria R. Capobianchi博士的基因组RNA。

CU 芯柱可实现细间距和高频...
引言 产业界要求器件薄、轻、短、小、性能高,细间距、高密度封装成为必然手段。然而,为了完全实现产业化,许多特性还有待改进,如散热、导电性、热导率、尺寸精度等。此外,在3D封装组装结构中,特别是像堆叠封装(PoP),焊料凸块可能会因为顶部封装的重量而坍塌。几年前,产业界引入了铜芯焊球来改善这些问题。顾名思义,铜芯焊球以球形铜为芯,在中心镀镍和焊料[1]-[2]。镀镍可有效防止锡和铜之间的扩散。铜芯焊球本身具有优异的导电特性和间隙高度优点,可以控制和保持一致的空间,防止封装之间的凸块坍塌。除此之外,Cu还有三大物理特性:高熔点(1083℃)、高电导率、高热导率。