XiaoMi-AI文件搜索系统
World File Search Systemgenomes

使用生成ai
癌症源自基因组的改变,了解这些变化如何导致疾病对于实现精确肿瘤学的目标至关重要。将基因组改变与健康结果联系起来需要使用准确的算法进行广泛的计算分析。多年来,这些算法已经变得越来越复杂,但是绝对的开放访问金标准数据集的严重短缺提出了一个基本挑战。由于基因组数据被视为个人健康信息,因此只能共享和重新分布一定数量的深入测序遗留癌症基因组。因此,工具基准测试通常是在与较旧技术和不确定基础真相的相同基因组集上进行的。这是开发改进分析工具的主要障碍。为了解决这个问题,我们开发了Oncogan,这是一种新型的生成AI工具,它结合了生成性对抗网络和表格变异自动编码器,以基于源自大规模基因组项目的训练集生成现实但完全合成的癌症基因组。我们的结果表明,这种方法准确地再现了多种常见癌症类型的体细胞突变,拷贝数改变和结构变异的规模,分布和特征,同时保护捐助者的隐私信息。Oncogan准确地概括了肿瘤类型特异性突变特征以及体细胞突变的位置分布。为了评估模拟的保真度,我们使用DeepTumour测试了合成基因组,该软件能够根据突变模式识别肿瘤类型,并证明了合成基因组肿瘤类型和DeepTumour类型的预测之间的一致性很高。我们还表明,使用Oncogan生成的合成数据增加实际供体数据可用于训练更准确的DeepTumour版本。

水平基因转移如何塑造昆虫基因组的进化?
昆虫是一个高度多样化的谱系,占所有描述的动物的50%,约有30个订单(Chapman,2009; Forister等,2019; Novotny等,2002)。昆虫是在大多数陆地和水生环境中发现的(Gullan&Cranston,2014; Scudder,2017),并且以多种方式成为生态系统健康的关键,例如通过充当分解,猎物,捕食者,捕食者和传粉者(Gurr等,2003; Majeed等,20222)。此外,它们相对较小的尺寸和高生殖率使它们能够占据大型生物所无法的多种生态壁ches(Berger等,2008; Gullan&Cranston,2014)。昆虫还与微型ISM(例如细菌和真菌)广泛相互作用,增强了昆虫适应不同环境的能力。微型肌肉是在昆虫的外骨骼,肠道和血液中以及内部昆虫细胞中发现的。昆虫肠道菌群有助于宿主的消化和
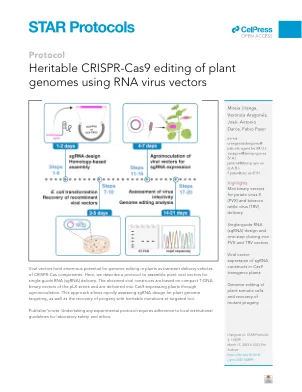
利用 RNA 病毒载体对植物基因组进行可遗传的 CRISPR-Cas9 编辑
病毒感染的系统性传播促进了编辑成分在植物组织内的积累。这导致了高效和快速的基因组编辑,从而为评估单向导 RNA (sgRNA) 设计的有效性和特异性提供了理想的筛选工具。几种基于植物 RNA 病毒的复制子已成功用于在组成性表达 Cas9 核酸酶的转基因植物中传递 sgRNA。9–19 然而,每种病毒载体都有自己的分子生物学特性,并且仅限于特定的宿主范围。在这里,我们描述了两种源自马铃薯病毒 X (PVX;Potexvirus 属) 和烟草脆裂病毒 (TRV;Tobravirus 属) 的病毒载体的工程改造,用于在模型物种本氏烟中传递非间隔 sgRNA(图 1)。所提出的 PVX 系统由单个二元载体 pLX-PVX 组成,该载体包含 PVX 基因组序列和一个来自竹花叶病毒 (BaMV) 的异源亚基因组启动子以驱动插入表达 (图 2)。TRV 系统依赖于 pLX-TRV1 和 pLX-TRV2,这是两个具有兼容来源的 T-DNA 载体,可同时进行病毒基因组成分的农杆菌接种 (JoinTRV)。pLX-TRV1 提供复制酶功能,而 pLX-TRV2 包含一个工程化的 TRV RNA2 序列和一个来自豌豆早褐病毒 (PEBV) 的异源亚基因组启动子以驱动插入表达 (图 2)。这两个病毒系统均基于 pLX 系列的紧凑 T-DNA 二元载体20,这些载体已成功用于通过农杆菌介导的接种 (农杆菌接种) 启动 RNA 和 DNA 病毒感染。 21–23 重组病毒复制子与 sgRNA 构建体组装并通过农杆菌接种递送到表达 Cas9 的植物中。系统性病毒感染导致生殖系基因组编辑和编辑后代的恢复(图 1)。

水平基因转移如何塑造昆虫基因组的进化?
昆虫是一个高度多样化的谱系,占所有描述的动物的50%,约有30个订单(Chapman,2009; Forister等,2019; Novotny等,2002)。昆虫是在大多数陆地和水生环境中发现的(Gullan&Cranston,2014; Scudder,2017),并且以多种方式成为生态系统健康的关键,例如通过充当分解,猎物,捕食者,捕食者和传粉者(Gurr等,2003; Majeed等,20222)。此外,它们相对较小的尺寸和高生殖率使它们能够占据大型生物所无法的多种生态壁ches(Berger等,2008; Gullan&Cranston,2014)。昆虫还与微型ISM(例如细菌和真菌)广泛相互作用,增强了昆虫适应不同环境的能力。微型肌肉是在昆虫的外骨骼,肠道和血液中以及内部昆虫细胞中发现的。昆虫肠道菌群有助于宿主的消化和

利用 RNA 病毒载体对植物基因组进行可遗传的 CRISPR-Cas9 编辑
液氮 n/an/a 关键商业检测 NEBuilder® HiFi DNA 组装预混液 New England Biolabs E2621S BsaIHF®v2 (20 U/µL) New England Biolabs R3733S Phusion TM 高保真 DNA 聚合酶 Thermo Fisher Scientific F530S MluI (10 U/µL) Thermo Fisher Scientific ER0561 ApaI (10 U/µL) Thermo Fisher Scientific ER1411 XhoI (10 U/µL) Thermo Fisher Scientific ER0691 EcoRI (10 U/µL) Thermo Fisher Scientific ER0271 RevertAid TM 逆转录酶 Thermo Fisher Scientific EP0441 RiboLock RNase 抑制剂 (40 U/μL) Thermo Fisher Scientific EO0381 NucleoSpin® 质粒试剂盒 Macherey-Nagel 740588.250 Zymoclean 凝胶 DNA 回收试剂盒 Zymo Research D4001 Zymo-Spin I Zymo Research C1003-250 嗜热菌 (Tth) DNA 聚合酶 Biotools 10.003 寡核苷酸 D1789 GGGAATCAATCACAGTGTTGGC

比较基因组学揭示了大规模混乱基因组的进化起源
摘要 纤毛虫是经历广泛程序性基因组重排的微生物真核生物,这是一种自然的基因组编辑过程,可将较长的生殖系染色体转换为较小的富含基因的体细胞染色体。三种研究较为深入的纤毛虫包括 Oxytricha trifallax 、 Tetrahymena thermophila 和 Paramecium tetraurelia ,但只有 Oxytricha 谱系具有大量乱序基因组,其在发育过程中的组装需要数十万个精确编程的 DNA 连接事件,代表了已知生物中最复杂的基因组动态。在这里,我们通过检查 Oxytricha 谱系中不连续和乱序基因的起源和进化来研究这种复杂基因组的出现。本研究比较了来自三个物种的六个基因组,即 Euplotes woodruffi、Tetmemena sp. 和模型纤毛虫 O. trifallax 的生殖系和体细胞基因组。我们对 E. woodruffi 的生殖系和体细胞基因组(它是一个外群)以及 Tetmemena sp 的生殖系基因组进行了测序、组装和注释。我们发现 Tetmemena 的生殖系基因组与 Oxytricha 的一样具有严重的杂乱和中断:13.6%的基因位点需要程序性易位和/或倒位,一些基因在发育过程中需要数百个精确的基因编辑事件。这项研究表明,早期分化的螺旋藻 E. woodruffi 也有一个杂乱的基因组,但只有大约一半的基因位点(7.3%)是杂乱的。此外,它的杂乱基因不太复杂,共同支持了 Euplotes 作为此谱系中可能的进化中间体的地位,处于积累复杂的进化基因组重排的过程中,所有这些都需要大量修复来组装功能性编码区。比较分析还表明,混乱的基因座通常与局部重复有关,支持了通过许多小的 DNA 重复和衰减事件来产生复杂的、混乱的基因组的渐进模型。

使用 QIAGEN ® CLC Genomics Workbench 组装和注释质体基因组
本应用说明介绍了使用 QIAGEN CLC Genomics Workbench 进行质体组装的三种不同工作流程。工具和工作流程的选择取决于目标物种中质体的结构以及测序数据的类型。组装具有长 IR 的质体需要足够长的读取以跨越重复。这种长读取通常保真度较低,组装需要完善。组装没有长 IR 的质体可以使用“较短”的高保真长读取来实现,并且不需要重叠群完善。我们强调的另一个步骤是在组装质体之前减少 NGS 数据集。我们描述了从全基因组测序数据中预选和不预选叶绿体读取的不同从头组装工作流程。

Panorama:探索种间多样性的微生物基因组的比较pangenomics工具
随着公共数据库中核基因组的增加,比较基因组学方法现在使用数百种基因组来分析物种多样性。许多研究着重于整个物种基因含量,即pangenome,以了解其在流行病学或环境数据方面的共同和可变基因方面的进化。在这种情况下,我们一直在研究基因组数据表示作为pangenome图。我们开发了用于重建和分配的pangenome重建和分区(Ppanggolin 1),基因组可塑性鉴定区域(PANRGP 2)和模块检测(PanModule 3)的方法。与Panorama一起,我们将实现新的方法论发展,以进行pangemenomes的比较研究。 将有助于研究细菌的适应潜力,并更好地了解微生物代谢多样性背后的进化动力学。与Panorama一起,我们将实现新的方法论发展,以进行pangemenomes的比较研究。将有助于研究细菌的适应潜力,并更好地了解微生物代谢多样性背后的进化动力学。

来自哈萨克斯坦的七种假发物种的叶绿体基因组的比较分析
杜松种类是杯形科中的灌木或树木,在森林生态系统中起着重要作用。在这项研究中,我们报告了在哈萨克斯坦收集的五种假发物种的质体(PT)基因组的完整序列(j。 communis,j。 Sibirica,J。 pseudosabina,j。 semiglobosa和j。 Davurica)。 除了两个完整的Pt基因组序列外,还注释了五种物种的Pt基因组的序列。 Sabina和J。 Seravschanica,我们先前已报告。 将这七种物种的Pt基因组序列与Pub-lic ncbi数据库中可用的杜松物种的Pt基因组进行了比较。 杜松物种的PT基因组的总长度,包括先前发表的PT基因组数据,范围为127,469 bp(j。 semiglobosa)至128,097 bp(j。 communis)。 每个杜松子质体由119个基因组成,包括82个蛋白质编码基因,33个转移RNA和4个核糖体RNA基因。 在确定的基因中,16个包含一个或两个内含子,并复制了2个tRNA基因。 对PT基因组序列的比较评估表明,鉴定了1145个简单序列重复标记。 基于82种蛋白质编码基因的26种假发物种的系统发育树,将杜松样品分为两个主要进化枝,对应于Juniperus和Sabina切片。 PT基因组序列的分析表明ACCD和YCF2是两个最多态性基因。在这项研究中,我们报告了在哈萨克斯坦收集的五种假发物种的质体(PT)基因组的完整序列(j。communis,j。Sibirica,J。 pseudosabina,j。 semiglobosa和j。 Davurica)。 除了两个完整的Pt基因组序列外,还注释了五种物种的Pt基因组的序列。 Sabina和J。 Seravschanica,我们先前已报告。 将这七种物种的Pt基因组序列与Pub-lic ncbi数据库中可用的杜松物种的Pt基因组进行了比较。 杜松物种的PT基因组的总长度,包括先前发表的PT基因组数据,范围为127,469 bp(j。 semiglobosa)至128,097 bp(j。 communis)。 每个杜松子质体由119个基因组成,包括82个蛋白质编码基因,33个转移RNA和4个核糖体RNA基因。 在确定的基因中,16个包含一个或两个内含子,并复制了2个tRNA基因。 对PT基因组序列的比较评估表明,鉴定了1145个简单序列重复标记。 基于82种蛋白质编码基因的26种假发物种的系统发育树,将杜松样品分为两个主要进化枝,对应于Juniperus和Sabina切片。 PT基因组序列的分析表明ACCD和YCF2是两个最多态性基因。Sibirica,J。pseudosabina,j。semiglobosa和j。Davurica)。 除了两个完整的Pt基因组序列外,还注释了五种物种的Pt基因组的序列。 Sabina和J。 Seravschanica,我们先前已报告。 将这七种物种的Pt基因组序列与Pub-lic ncbi数据库中可用的杜松物种的Pt基因组进行了比较。 杜松物种的PT基因组的总长度,包括先前发表的PT基因组数据,范围为127,469 bp(j。 semiglobosa)至128,097 bp(j。 communis)。 每个杜松子质体由119个基因组成,包括82个蛋白质编码基因,33个转移RNA和4个核糖体RNA基因。 在确定的基因中,16个包含一个或两个内含子,并复制了2个tRNA基因。 对PT基因组序列的比较评估表明,鉴定了1145个简单序列重复标记。 基于82种蛋白质编码基因的26种假发物种的系统发育树,将杜松样品分为两个主要进化枝,对应于Juniperus和Sabina切片。 PT基因组序列的分析表明ACCD和YCF2是两个最多态性基因。Davurica)。除了两个完整的Pt基因组序列外,还注释了五种物种的Pt基因组的序列。Sabina和J。Seravschanica,我们先前已报告。将这七种物种的Pt基因组序列与Pub-lic ncbi数据库中可用的杜松物种的Pt基因组进行了比较。杜松物种的PT基因组的总长度,包括先前发表的PT基因组数据,范围为127,469 bp(j。semiglobosa)至128,097 bp(j。communis)。每个杜松子质体由119个基因组成,包括82个蛋白质编码基因,33个转移RNA和4个核糖体RNA基因。在确定的基因中,16个包含一个或两个内含子,并复制了2个tRNA基因。对PT基因组序列的比较评估表明,鉴定了1145个简单序列重复标记。基于82种蛋白质编码基因的26种假发物种的系统发育树,将杜松样品分为两个主要进化枝,对应于Juniperus和Sabina切片。PT基因组序列的分析表明ACCD和YCF2是两个最多态性基因。使用这两个基因对26种假发物种的系统发育评估证实,它们可以有效地用作该属中植物分析的DNA条形码。这些假发物种的测序质体提供了大量遗传数据,这些数据对于该属的将来的基因组研究很有价值。

分析来自祖先多样化亚洲基因组的临床相关变体
©2022作者。本文是根据Creative Commons Attribution 4.0 International许可证的许可,该许可允许使用,共享,适应,分发和复制任何媒介或格式,因为您对原始作者和来源提供了适当的信誉,并提供了与Creative Commons许可证的链接,并指出是否进行了更改。本文章中的图像或其他第三方材料包含在文章的Creative Commons许可中,除非在材料的信用额度中另有说明。如果材料未包含在文章的Creative Commons许可中,并且您的预期用途不受法定法规的允许或超过允许的用途,则您将需要直接从版权所有者那里获得许可。要查看此许可证的副本,请访问http://creativecommons.org/licenses/4.0/。