XiaoMi-AI文件搜索系统
World File Search Systemmutation

CRISPR-CAS9介导的甲基转移酶METTL4突变导致胚胎bbybyx mori
摘要:最近发现DNA N6-甲基趋化(6MA)在基因中扮演调节作用,该作用与真核物种的各种生物学过程联系起来。6MA甲基转移酶的功能鉴定对于理解表观遗传6MA甲基化的潜在分子机制至关重要。据报道,甲基转移酶METTL4可以催化6ma的甲基化。但是,METTL4的功能在很大程度上未知。在这项研究中,我们旨在研究Bombyx Mori同源性METTL4(BMMETTL4)在鳞翅目模型昆虫中的作用。通过使用CRISPR-CAS9系统,我们在蚕中对BMMETTL4进行了体积突变,发现BMMETTL4的破坏会导致蚕胚晚期的发育缺陷和随后的致死性。我们进行了RNA-Seq,并确定了BMMETTL4突变体中有3192个差异表达的基因,其中包括1743个上调和1449个下调的基因。基因和基因组分析的基因本体论和京都百科全书表明,涉及分子结构,几丁质结合和丝氨酸水解酶活性的基因受BMMETTL4突变的显着影响。我们进一步发现,表皮蛋白基因和胶原蛋白的表达明显降低,而胶原酶高度增加,这对异常的胚胎和蚕的孵化性降低了。采取了这些结果,这些结果表明6MA甲基转移酶BMMETTL4在调节蚕的胚胎发育中的关键作用。

肥厚型心肌病大鼠模型中 MYBPC3 基因突变的同源定向修复
对照 1098hom 大鼠的心脏功能受损。对照 1098hom 大鼠的射血分数 (EF) 为 61%,而 WT 组为 78%。对照 1098hom 大鼠的缩短分数 (FS)、最大 dP/dt 和最小 dP/dt 也下降。HDR 治疗部分恢复了受损的心脏功能(图 4a、b、c、d 和表 S2)。此外,心房利钠肽 (ANP) 和脑利钠肽 (BNP) 是心力衰竭的生物标志物,在对照 1098hom 大鼠心脏中升高,通过 HDR 编辑标准化(图 4e、f)。心脏切片的天狼星红染色显示对照 1098hom 大鼠的心脏纤维化增加,HDR 治疗可减轻这种纤维化。有无对照的 1098hom 大鼠的心脏肥大和心脏功能没有显着差异

基因组编辑模拟和逆转与骨髓增生性肿瘤相关的普遍突变
骨髓增生性肿瘤 (MPN) 会导致血细胞(如红细胞增多症)或血小板(原发性血小板增多症)的过度生成。JAK2 V617F 是许多 MPN 中最常见的体细胞突变,但之前在小鼠中对这种突变的建模依赖于转基因过度表达,并导致不同的表型,在某些情况下,这些表型归因于表达水平。CRISPR-Cas9 工程通过精确修改原代细胞中的内源性位点,为建模和潜在治愈遗传编码疾病提供了新的可能性。我们在此开发了“无疤痕”的 Cas9 试剂,用于在永生化人类红系祖细胞 (HUDEP-2)、CD34+ 成人人类造血干细胞和祖细胞 (HSPC) 以及免疫表型长期造血干细胞 (LT-HSC) 中创建和逆转 JAK2 V617F 突变。我们发现与内源性 JAK2 V617F 等位基因相关的体外增殖没有明显增加,但与野生型细胞共培养揭示了突变提供的竞争性生长优势。即使在没有造血细胞因子信号传导的情况下,获得 V617F 等位基因也会促进红系祖细胞的终末分化。综上所述,这些数据与 MPN 的逐渐进展的表现相一致,并表明与转基因过表达模型相比,内源性获得性 JAK2 V617F 突变可能产生更细微的表型。

SPEG中的一种新型隐性突变导致早期发作扩张性心肌病
1索罗卡大学医学中心儿科心脏病学系和卫生科学学院,本盖尔本·尼加大学,以色列比尔 - 肖瓦,2什拉加·西格尔(Shraga Segal Segal)微生物学,免疫学和遗传学系,伊格里恩·尼加(Negev)内加尔本盖尔大学内盖伊大学,以色列比比尔·肖瓦大学,4号心胸外科系,斯坦福大学医学院,美国加利福尼亚州斯坦福大学医学院,美国,美国斯坦福大学5心血管研究所,斯坦福大学医学院,斯坦福大学斯坦福大学,加利福尼亚州斯坦福大学,美国加利福尼亚州,美国6个Cardiovel Instuctive and Stanford Instervation of Stanford and Stanford and Stanford Satnford of Stanforf美国加利福尼亚州斯坦福大学的斯坦福大学眼科与医学学院,美国尼加夫妇本·古里安大学,啤酒士本盖尔大学,再生医学和干细胞研究中心,以色列9

酿酒酵母基因组必需基因的有效靶向突变
摘要 通过等位基因置换进行基因定点突变是功能基因组分析和代谢工程的重要内容,但传统的通过选择标记对必需基因进行定点突变的方法存在很大挑战,因为第一步必需基因敲除将导致致死的表型。本文利用CRISPR/Cas9系统,发展了一种两端选择标记(Two-ESM)方法对酿酒酵母中的必需基因进行定点突变,成功构建了酿酒酵母必需基因ERG20(编码法呢基二磷酸合酶)的单突变和双突变体,突变效率达100%。此外,与传统方法相比,Two-ESM方法显著提高了突变效率,简化了遗传操作程序。通过动态调控突变基因的表达和整合模块的优化,进一步提高了基因组整合和突变效率。这种 Two-ESM 方法将有助于构建酵母功能基因组分析和代谢通量调控所需的必需基因的基因组突变。

早期小脑发育的调节
单个细胞的线粒体DNA(mtDNA)的测序已在本地人类样品和临床标本中解析了克隆性和谱系。先前的工作表明,杂质mtDNA变体可用于描述造血的克隆性,但它们重建细胞系统发育的能力有限。但是,Weng等人的最新报告。 通过描述据报道解决高分辨率系统发育树的细胞之间的空前数量的共享mtDNA变体来挑战当前范式。 我们重新审查了Weng等人的主张,并确定了有关这种前所未有的联系的两个主要关注点。 首先,在每个细胞的单个分子中检测到细胞之间的共享变体,其次,这些变体在mtDNA分子的边缘富集了10-20倍,使人联想到其他测序方法中报道的文物。 此外,我们的分析表明,修剪低支撑和可能的人造mtDNA变体几乎消除了所有报道的系统发育结构。 因此,我们强烈警告不要使用依赖最小证据的mtDNA变体工作流,包括Weng等人在Weng等人中引入的计算管道,因为具有高连接性和较低证据的变体可能是导致假系统发电的构建的伪像。但是,Weng等人的最新报告。通过描述据报道解决高分辨率系统发育树的细胞之间的空前数量的共享mtDNA变体来挑战当前范式。我们重新审查了Weng等人的主张,并确定了有关这种前所未有的联系的两个主要关注点。首先,在每个细胞的单个分子中检测到细胞之间的共享变体,其次,这些变体在mtDNA分子的边缘富集了10-20倍,使人联想到其他测序方法中报道的文物。此外,我们的分析表明,修剪低支撑和可能的人造mtDNA变体几乎消除了所有报道的系统发育结构。因此,我们强烈警告不要使用依赖最小证据的mtDNA变体工作流,包括Weng等人在Weng等人中引入的计算管道,因为具有高连接性和较低证据的变体可能是导致假系统发电的构建的伪像。

初级保健中触发工具的开发和验证
氨基酰基-TRNA和GTP结合的翻译伸长因子EF-TU识别核糖体的A位点密码子取决于多肽(P)和出口(E)密码子位点中存在的密码子和TRNA物种。为了了解密码子环境如何影响tRNA结合的EF-TU识别密码子识别的效率,开发了一个遗传系统,可以通过慢速翻译密码子组合选择快速翻译。选择通过慢速翻译的UCA-UAC对,两侧是Histi Dine密码子,从而在必需的TRNA Leuz的D-STEM中分离了A25G碱基取代突变体,该突变体识别UUA和UUG亮氨酸密码子。Leuz(A25G)替换允许通过包括UCA密码子在内的所有密码子对进行更快的翻译。插入。这项工作是根据trpt tRNA中的Hirsh UGA非理性抑制剂G24a突变所做的,它提供了遗传证据,即通过伸长因子TU进行的GTP后水解校对校验拟合步骤可以通过TRNA物种铰链区域中的结构相互作用来控制。我们的结果支持一个模型,在该模型中,mRNA翻译中的tRNA弯曲成分允许EF TU时间增强其区分cognate和接近同名mRNA密码子之间的tRNA相互作用的能力。
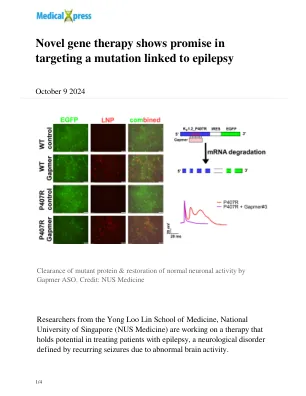
新型基因疗法在靶向与癫痫有关的突变
一种称为Gapmer反义寡核苷酸(ASO)的专门治疗方法旨在专门靶向和分解故障的核糖核酸(RNA),同时保持正常基因功能完整。使用这种RNA疗法导致在KCNA2基因中编码的有问题的钾通道蛋白中显着降低,这有助于恢复正常的钾流量并减少与癫痫有关的过度神经元活性。

一种遗传的mtDNA突变重塑巨噬细胞和体内炎症细胞因子反应
巨噬细胞中线粒体生物能的受损可能会驱动高炎性细胞因子反应1-6,但是是否也可能是由遗传的mtDNA突变引起的。在这里,我们使用一种多摩变方法来解决这个问题,该方法将超分辨率成像和代谢分析整合到来自丙氨酸7的线粒体trNA中异质质突变(M.5019a> g)的线粒体疾病的小鼠模型中的巨噬细胞。这些M.5019a> G巨噬细胞在呼吸链复合物中表现出缺陷,并且由于中骨内部翻译减少而导致氧化磷酸化(OXPHOS)。以适应这种代谢应激,线粒体融合,还原性谷氨酰胺代谢和有氧糖酵解均增加。在炎症激活后,I型干扰素(IFN-I)释放得到增强,而在M.5019a> G巨噬细胞中限制了促炎性细胞因子和黄磷脂的产生。最后,使用M.5019a> G小鼠的体内内毒素性模型显示IFN-I水平和疾病行为升高。总而言之,我们的研究确定了响应致病性mtDNA突变的先天免疫信号传导的意外失衡,对MTDNA疾病患者的病理发展具有重要意义。8。

SLC4A10突变引起与GABA能传播受损相关的神经系统疾病
SLC4A10是一种血浆膜结合的转运蛋白,它利用Na +梯度驱动细胞HCO 3-摄取,从而介导酸挤出。在哺乳动物大脑中,SLC4A10在主要神经元和中间神经元以及脉络丛的上皮细胞中表达,该器官调节CSF的产生。使用五个无关家庭的样本中的下一代测序,包括九个受影响的个体,我们表明双重性SLC4A10功能丧失变体会导致人类临床上可识别的神经发育障碍。该病情的基本临床特征包括婴儿期肌张力障碍,所有领域的精神运动延迟发展和智力障碍。受影响的个体通常显示出与自闭症谱系障碍有关的特征,包括焦虑,多动症和刻板动作。在两种情况下,据报道,在生命的最初几年中,癫痫发作的发作是分离的,进一步影响的儿童在没有明显的临床癫痫发作的情况下在脑电图上表现出了暂时性的癫痫发作。据报道枕骨围在出生时正常,但在10个受影响的个体中,有7个进化了出生后的小头畸形。神经放射学特征包括与枕骨圆周相比的相对保留,特征性狭窄有时“裂开”的侧脑室和call体异常。SLC4A10 - / - 小鼠,缺乏SLC4A10,还显示出小的侧脑室和轻度的行为异常,包括延迟的习惯和两目标新颖对象识别任务的改变。在SLC4A10 - / - 小鼠和受影响的个体中崩溃的脑腹膜cles cles表明SLC4A10在CSF的生产中起着重要作用。然而,值得注意的是,尽管CSF在发育中的大脑和成年大脑中的各种作用,但SLC4A10 - / - 小鼠的皮质似乎非常完整。与突触标记的共同染色表明,在神经元中,SLC4A10定位于抑制性,但不能兴奋性的午睡。这些发现得到了我们的功能研究的支持,该研究表明,在SLC4A10 - / - 小鼠中释放了抑制性神经肌群的释放,而兴奋性神经递质谷氨酸的释放则保留了。对细胞内pH的操纵部分挽救了GABA释放。我们的研究共同定义了一种与SLC4A10中双重性致病变异相关的新型神经发育障碍,并强调了SLC4A10功能丧失对脑发育,突触传播和网络特性的进一步分析的重要性。