XiaoMi-AI文件搜索系统
World File Search System变构

建立 CRISPR-Cas9 中的变构机制
变构是蛋白质的基本特性,它调节空间上相距遥远的位点之间的生化信息传递。在这里,我们报告了分子动力学 (MD) 模拟在发现 CRISPR-Cas9 中的变构通讯机制方面的关键作用,CRISPR-Cas9 是一种领先的基因组编辑机制,在医学和生物技术方面具有巨大的前景。MD 揭示了变构如何在 CRISPR-Cas9 功能的至少三个步骤中发挥作用:影响 DNA 识别、介导切割和干扰脱靶活性。发现激活协同 DNA 切割的变构通讯通过连接 HNH 和 RuvC 催化域的 L1/L2 环进行。这些“变构传感器”的识别启发了具有改进特异性的 Cas9 蛋白新变体的开发,为控制 CRISPR-Cas9 活性开辟了一条新途径。讨论的研究还强调了识别叶在催化 HNH 域的构象激活中的关键作用。具体而言,REC3 区域被发现通过感知 RNA:DNA 杂合体的形成来调节 HNH 的动态。REC3 的作用在 DNA 错配的情况下尤其重要。事实上,REC3 对在特定位置含有错配对的 RNA:DNA 杂合体的干扰导致 HNH 锁定在非活性“构象检查点”构象中,从而阻碍脱靶切割。总体而言,MD 模拟建立了 CRISPR-Cas9 变构现象的基本机制,有助于开发新的 CRISPR-Cas9 变体以改进基因组编辑的工程策略。

KRAS抑制的能量和变构景观
这是接受出版的同行评审纸的PDF文件。尽管未经编辑,但内容已受到初步格式。自然正在为排版纸的早期版本作为我们的作者和读者的服务。文本和数字将在本文以最终形式发表之前进行复制和证明审查。请注意,在生产过程中可能会发现可能影响内容的错误,并且所有法律免责声明都适用。
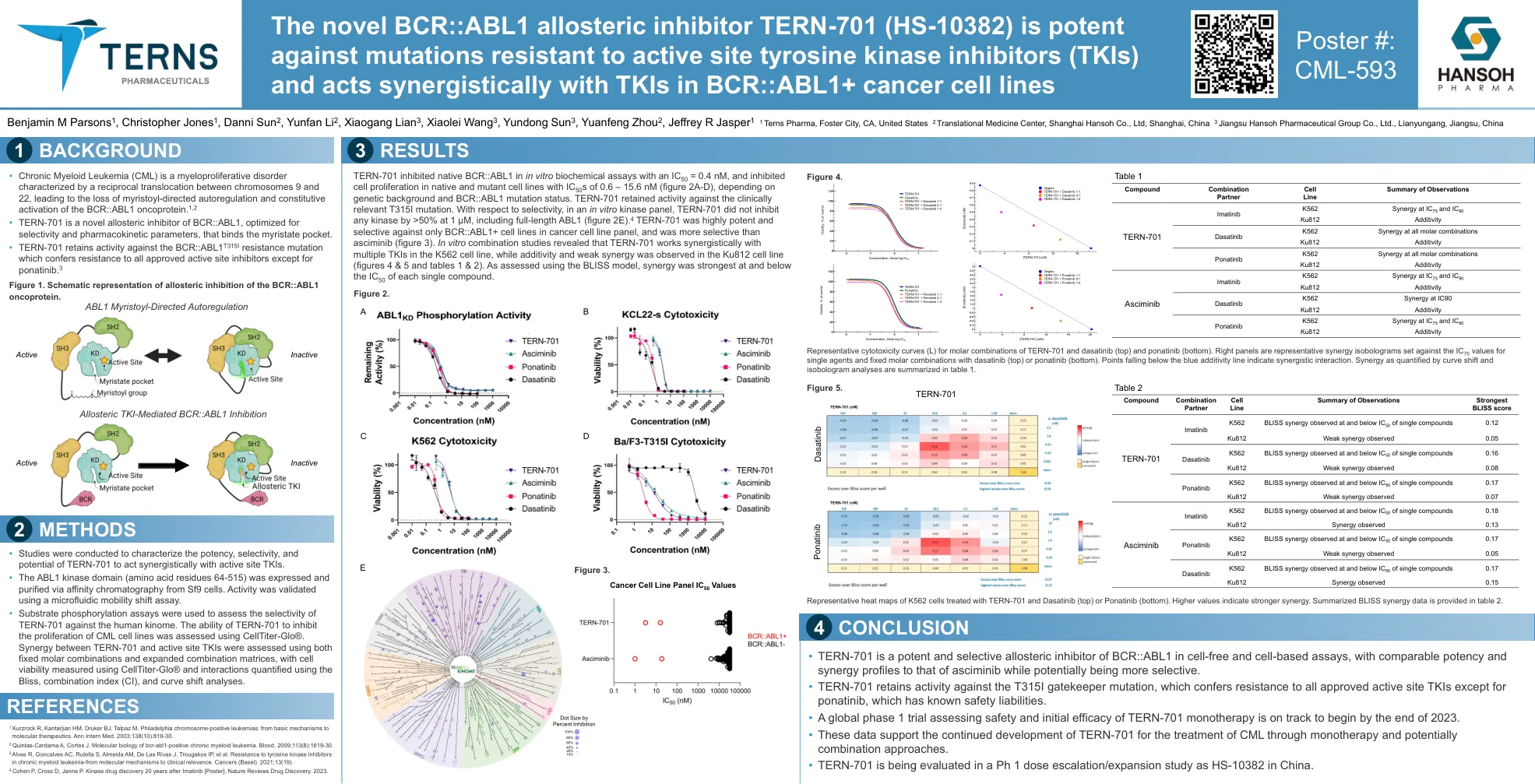
ABL1变构抑制剂Tern-701(HS-10382)有效
•进行研究以表征Tern-701与主动位点TKIS协同作用的效力,选择性和潜力。•ABL1激酶结构域(氨基酸残基64-515)通过SF9细胞的亲和力色谱法表达并纯化。使用微流动迁移率转移测定法对活性进行了验证。•使用底物磷酸化测定法来评估tern-701对人类组的选择性。使用CellTiter-Glo®评估了Tern-701抑制CML细胞系增殖的能力。使用固定摩尔组合和扩展的组合矩阵评估了Tern-701和主动位点TKI之间的协同作用,并使用CellTiter-Glo®测量细胞活力,并使用Bliss,组合指数(CI)和曲线移位分析进行了量化的相互作用。

解码变构景观-RSC Publishing
和跟踪原子运动,从而提供了详细的见解,对构象变化和分子动力学。5通常,MD模拟从实验确定的三维结构开始,随后能量最小化和对近似生理条件的平衡。MD模拟的强度在于它们能够揭示各个时间尺度上符合符合性变化的能力,从而提供了动态的信息,这很难通过传统的实验方法获得,尤其是在酶变构调节的背景下。变构调节是指通过构象变化调节酶活性的过程,通常会参与关键分子间相互作用的动态调整。由于这些过渡发生在次纳秒至millise-cond时标,因此他们具有挑战性地使用传统的实验技术直接观察。MD模拟提供了很高的时间分辨率,从而实现了调节机制的表征。通过跟踪酶构象变化和内部分子动力学,MD模拟有助于鉴定控制酶活性和信号转导的变构位点,这通常是从单独静态结构分析中获得的信息。6

STX-478,一种突变选择性变构 PI3Kα 抑制剂...
磷酸肌醇 3-激酶 (PI3K)/AKT 轴在癌症发展中起着关键作用 (1, 2),约 14% 的癌症 (3, 4) 中发生 PI3K α 致癌基因变异。这些突变遍布整个 PIK3CA 基因,但在螺旋 (E542K、E545K) 和激酶 (H1047R/L) 结构域的热点氨基酸位点高度富集 (3, 5),尽管 PI3K α 热点突变的频率因癌症而异 (6)。PI3K α 突变在乳腺癌中最为常见,发生在约 36% 的患者中,其中约 28% 为螺旋结构域突变,40% 为激酶结构域突变 (7)。突变型 PI3K α 也是其他难治性癌症的常见致癌驱动因素,包括胃癌 (15%)、结肠癌 (25%)、头颈部鳞状细胞癌 (HNSCC;13%) 和子宫癌 (45%;参考文献 3、8、9)。 PI3K α 异构体选择性抑制的临床益处已在 PI3K α 突变型癌症中得到证实。 Alpelisib 是一种正构抑制剂,对野生型 (WT) 和突变型 PI3K α 均具有同等抑制作用。

Sirtuin 2脱乙酰基酶活性的变构抑制剂...
不应发展抵抗。此外,由于不同的病毒使用重叠的细胞途径和因素来支持其复制(4)和抗病毒防御系统通常以这些常见途径为目标,因此HTA可以表现出广泛的光谱活性(5)。因此,HTA具有治疗病毒疾病的类别,而病毒剂跨越了多个病毒家族。重要的是,广谱HTA具有在大流行时提供快速治疗溶液的潜力,从而减少了新型病毒鉴定和药理干预之间的时间(6,7)。超出了这种定期需求,HTA可以治疗患有不同家族病毒感染风险的患者,例如在免疫抑制治疗期间患有疱疹病毒,帕托病毒,多瘤病毒,肝瘤病毒,肝癌,肝癌和可可菌感染风险升高的患者(8、9)。

纳米构造对人ABCG2的变构抑制的结构基础
ABCG2是一种ATP结合盒转运蛋白,它导出了多种异种生物化合物,并被认为是癌细胞中多药耐药性的因素。底物和与ABCG2的相互作用进行了广泛的研究,并且已经开发出了小分子抑制剂,以防止从肿瘤细胞中输出抗癌药物。在这里,我们探索了靶点位点以外的抑制剂的潜力。我们开发了针对ABCG2的新型纳米化,并使用功能分析选择了三种抑制性纳米型(NB8,NB17和NB96),通过单个粒子冷冻电子显微镜进行结构研究。我们的结果表明,这些纳米结合在变构与核苷酸结合域的不同区域结合。NB8的两个副本与NBD的顶点结合,以防止它们完全关闭。NB17在转运蛋白的两倍轴附近结合,并与两个NBD相互作用。NB96与NBD的侧面结合,并固定与与ATP结合和水解相关的关键基序连接的区域。所有三种纳米体都阻止了转移者经历底物运输所需的构象变化。这些发现提高了我们对外部粘合剂调节ABCG2的分子基础的理解,这可能会促进新一代抑制剂的发展。此外,这是通过纳米剂对人多药耐药转运蛋白进行调节的第一个例子。2023作者。由Elsevier Ltd.这是CC下的开放式访问文章(http://creativecom- mons.org/licenses/4.0/)。

多级力依赖性变构增强Ae-
经典的钙粘蛋白是跨膜蛋白,其细胞外域连接相邻细胞,其细胞内结构域通过B-蛋白酶和A -Catenin连接到肌动蛋白细胞骨架。cadherin- catenin络合物传递了驱动组织形态发生和伤口愈合的力。此外,E-catenin构象的张力依赖性变化使其能够募集肌动蛋白结合蛋白葡萄蛋白到细胞 - 细胞连接蛋白,从而有助于连接性增强。多种钙粘蛋白复合物的方式以及是否合作以加强对负载的细胞 - 细胞连接的构成。在这里,我们使用了单分子光学陷阱测量值来检查多种钙粘蛋白 - catenin络合物如何在负载下与F-肌动蛋白相互作用,以及这种相互作用如何受到杂质蛋白的影响。我们表明,朝向肌动灯的()末端的力导致平均寿命长3倍,比将力施加到刺(+)末端时。我们还通过包含钙粘蛋白 - 钙蛋白复合物和葡萄蛋白头部区域的第四纪复合物测量了依赖性的肌动蛋白结合,它们本身无法结合肌动蛋白。该四元复合物的结合寿命随着额外的配合物结合的F-肌动蛋白而增加,但仅当载荷朝向()末端定向时。相比之下,单独的钙粘蛋白 - 钙蛋白复合物并未显示这种合作的形式。2023 Elsevier Ltd.保留所有权利。这些发现揭示了多级,力依赖性调节,从而增强了多个钙粘蛋白/catenin络合物与F-肌动蛋白的缔合的强度,从而提供了阳性反馈,从而可以增强结并促进F-肌动蛋白,从而促进高阶细胞骨架组织的出现。

基于肽的变构抑制剂靶向 TNFR1 ... - DR-NTU
Chih Hung Lo 1,#, * 1 Lee Kong Chian School of Medicine, Nanyang Technological University, Singapore 308232, Singapore 2 School of Applied Science, Republic Polytechnic, Singapore 738964, Singapore 3 Centro Multidisciplinario de Estudios en Biotecnología, Facultad de Medicina Veterinaria y Zootecnia, Universidad Michoacana de San Nicolás de Hidalgo, Morelia 58893, México 4 School of Chemical Sciences, Meritorious Autonomous University of Puebla (BUAP), University City, Puebla 72570, México # 同等贡献 *通讯作者:Chih Hung Lo,博士 (chihhung.lo@ntu.edu.sg) Víctor M. Baizabal-Aguirre,博士 (victor.baizabal@umich.mx)关键词 TNFR1 信号传导,受体特异性抑制、构象动力学、非竞争性抑制、变构机制、药物发现、肽抑制剂、抗炎摘要肿瘤坏死因子 (TNF) 受体 1 (TNFR1) 在介导 TNF 诱导的信号通路和调节炎症反应中起关键作用。最近的研究表明,TNFR1 活化涉及配体前组装受体二聚体的构象重排,而靶向受体构象动力学是调节 TNFR1 信号的可行策略。在这里,我们结合使用生物物理、生化和细胞分析以及分子动力学模拟来表明抗炎肽 (FKCRRWQWRMKK)(我们称之为 FKC)通过改变受体二聚体的构象状态来变构抑制 TNFR1 活化,而不会阻断受体-配体相互作用或破坏受体二聚化。我们还通过展示该肽抑制 HEK293 细胞中的 TNFR1 信号传导并减轻腹膜内 TNF 注射小鼠的炎症来证明 FKC 的功效。从机制上讲,我们发现 FKC 与 TNFR1 富含半胱氨酸的结构域 (CRD2/3) 结合并扰乱受体激活所需的构象动力学。重要的是,FKC 增加了受体二聚体中 CRD2/3 和 CRD4 的开放频率,并诱导受体胞质区域的构象开放。这会导致抑制构象状态,阻碍下游信号分子的募集。总之,这些数据为靶向 TNFR1 构象活性区域的可行性提供了证据,并为受体特异性抑制 TNFR1 信号传导开辟了新途径。意义

基于DNAZyMe的可编程变构策略的DNA有限状态机
摘要:活生物体可以通过响应外部和内部刺激来产生相应的功能,而这种易怒性在自然界中起着关键作用。受到这种自然时间响应的启发,具有处理时间相关信息的纳米版本的开发和设计可以促进分子信息处理系统的开发。在这里,我们提出了一台可以动态响应顺序刺激信号的DNA有限状态机。为了构建这款状态机,开发了一种可编程的变构策略。该策略使用可构造的DNA发夹对DNAZYME构象进行可编程控制。基于此策略,我们首先实施了具有两个状态的有限状态机器。通过策略的模块化设计,我们进一步实现了具有五个状态的有限状态机器。DNA有限状态机赋予分子信息系统具有可逆逻辑控制和秩序检测的能力,可以将其扩展到更复杂的DNA计算和纳米机械,以促进动态纳米技术的发展。