XiaoMi-AI文件搜索系统
World File Search System宿主

寄生虫生存与宿主免疫之间的战争拖船
寄生疾病在热带和亚热带国家造成重大伤害,导致死亡率,发病率和社会经济差异。感染的结果和疾病的严重程度通常取决于寄生虫生存策略与宿主免疫反应之间的相互作用。强大而有效的宿主免疫反应可以限制寄生虫的复制,降低疾病的严重程度并促进康复。相反,如果寄生虫可以逃避或抑制免疫反应,则感染可能会持续并导致慢性或严重疾病。有时,宿主免疫反应本身可以通过寄生虫感染引发的过度或失调的免疫反应会导致疾病病理,从而导致组织损伤,炎症和免疫介导的疾病。寄生虫和宿主免疫之间的这场战争是由协同进化动力学驱动的。寄生虫不断适应托管免疫反应,然后托管又制定了应对寄生虫逃避策略的新策略。这种共同进化过程可以导致具有增强的免疫逃避能力(从而促进耐药性)和宿主遗传变异的新寄生虫菌株的出现,从而赋予耐药性或对特定寄生虫的敏感性。因此,了解这场正在进行的战斗中涉及的复杂机制和动态对于开发更有效的方法来管理和打击寄生疾病至关重要。它涉及研究寄生虫采用的机制,以逃避免疫检测和开发可以增强宿主免疫反应的干预措施。Nguyen等人的研究论文。此外,探索宿主对寄生虫感染的遗传因素的遗传因素可以帮助识别较高风险的人并为有针对性的预防或治疗方法提供信息。该研究主题旨在为我们的理解 - 寄生虫相互作用做出贡献,从而导致寄生虫学领域的进步和人类健康的改善。它包括四个原始研究文章和一篇评论文章,重点介绍锥虫病,利什曼病,疟疾,贾第鞭毛病毒(原生动物感染)和血吸虫病(蠕虫感染)。重点是由细胞外原生动物寄生虫锥虫埃文西(Trypanosoma evansi)引起的感染,该寄生虫在牲畜和游戏动物中导致致命的慢性浪费疾病。这项研究已建立了一个实验疾病模型

工厂Holobiont内的共同进化驱动宿主性能
植物与影响其生长和韧性的各种微生物相互作用,因此可以将其视为生态实体,即“植物holobionts”,而不是奇异生物。在植物霍洛比特(Holobiont)中,地下微生物群的组装由宿主,微生物和环境因素统治。在微生物感知下,植物会导致免疫信号传导,导致调节微生物群组成的因子分泌。此外,微生物之间的代谢相互作用和对抗是社区集会的驱动力。我们认为,在回避过程中选择了复杂的植物 - 微生物和微生物间相互作用,并可能促进植物及其相关微生物的生存和适应性作为Holobionts。作为此过程的一部分,植物发展了代谢物介导的策略,以选择性地募集其微生物群中的有益微生物。这些微生物群中的一些成员显示出宿主适应,互惠可能会迅速产生。在Holobiont中,微生物群也共同进化的拮抗活性,限制了具有高病原潜力的微生物的增殖,因此可以防止疾病发育。Holobionts内的共同进化最终会驱动植物的性能。

机器学习揭示了生产宿主链霉菌Albidoflavus
链霉菌Albidoflavus是一种流行且遗传上的平台菌株,用于通过异源生物合成基因簇(BGC)的表达进行自然产物发现和生产。然而,其转录调节网络(TRN)及其对继发代谢的影响尚不清楚。在这里,我们通过将独立的组件分析应用于来自内部和公共资源的218个高质量RNA-SEQ转录组的纲要,通过将独立的组件分析应用于88个独特的增长条件,来表征其TRN。我们获得了78个独立调制的基因集(imodulons),这些基因(imodulons)在定量地描述了跨不同条件的TRN及其活性状态。Through analyses of condition-dependent TRN activity states, we (i) describe how the TRN adapts to different growth conditions, (ii) conduct a cross-species iModulon comparison, uncovering shared features and unique characteristics of the TRN across lineages, (iii) detail the transcriptional activation of several endogenous BGCs, including surugamide, minimycin and paulomycin, and (iv) infer potential functions of 40% albidoflavus基因组中未表征的基因。我们的发现提供了对Albidoflavus的TRN的全面和定量的理解,为进一步的探索和实验验证提供了知识库。

感染沙门氏菌的宿主细胞的先天免疫反应
沙门氏菌是肠杆菌科家族中一种革兰氏阴性疗法的厌氧菌细菌,是肠道中普遍的病原体。沙门氏菌属内有两个不同的物种,即沙门氏菌肠和沙门氏菌,有超过2600多种由鞭毛和脂多糖抗原的变化区分的血清型。[1]。该细菌是一种人畜共患病原体,具有广泛的宿主,可以通过多种导致疾病的途径传播。在美国,每年有超过900万例食物和水生病病例,细菌造成了39%的这些病例[2]。非细类沙门氏菌占此类病例的30%,使其成为仅次于诺如病毒的粮食源性疾病的第二大细菌原因[2]。在用沙门氏菌感染后,宿主将触发其先天免疫反应。先天免疫系统由各种成分组成,例如先天免疫识别受体,细胞因子和先天免疫细胞,它们共同限制病原体感染并消除入侵的病原体。当病原体成功穿透宿主的物理屏障时,先天免疫系统首先使用模式识别受体(PRR)来识别病原体相关的分子模式(PAMP)并识别病原体[3]。然后,它启动了一系列防御机制,以快速反应并消除病原体。上皮细胞层形成了先天免疫系统的主要保护屏障,并具有粘液,抗菌剂和肠道微生物群,共同努力阻止病原体的侵袭[4]。还参与了对沙门氏菌的先天免疫反应[5]。本评论的重点是沙门氏菌感染的过程和宿主先天

功能性肿瘤细胞-内在 STING,而非宿主...
/-) 小鼠来评估局部肿瘤变化、肺转移、对远处肿瘤的远隔效应以及肿瘤微环境 (TME) 中的免疫细胞动态。结果结果表明,冷冻消融疗效依赖于适应性免疫和 STING 信号通路。与当前文献认为宿主衍生的 STING 激活作为体内抗肿瘤免疫驱动因素的重要作用相反,我们表明局部肿瘤控制、肺转移和对远处肿瘤的远隔效应都关键依赖于功能性的肿瘤细胞内在 STING 信号通路,该通路在冷冻消融的 TME 中诱导炎症趋化因子和细胞因子反应。这种依赖不仅限于冷冻消融,还包括肿瘤内 STING 激动剂治疗。此外,对临床肿瘤样本的基因表达数据库和组织微阵列的调查揭示了 STING 相关信号成分的广泛表达。结论肿瘤细胞内在 STING 通路是冷冻消融有效性的关键组成部分,表明 STING 相关信号成分的表达可作为潜在的治疗反应生物标志物。我们的数据还强调迫切需要进一步表征肿瘤细胞内在 STING 通路以及冷冻消融和其他 STING 依赖性治疗方法引起的相关下游炎症反应。
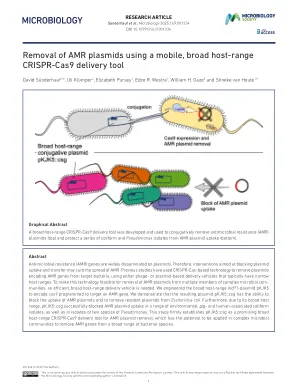
使用移动、广泛的宿主去除 AMR 质粒
抗菌素耐药性 (AMR) 基因广泛传播于质粒上。因此,旨在阻断质粒吸收和转移的干预措施可能会抑制 AMR 的传播。先前的研究已使用基于 CRISPR-Cas 的技术从目标细菌中去除编码 AMR 基因的质粒,使用通常宿主范围较窄的噬菌体或质粒递送载体。为了使该技术可用于从复杂微生物群落的多个成员中去除 AMR 质粒,需要一种高效、宿主范围广的递送载体。我们设计了宿主范围广的 IncP1 质粒 pKJK5 来编码被编程为靶向 AMR 基因的 cas9。我们证明所得质粒 pKJK5::csg 能够阻断 AMR 质粒的吸收并去除大肠杆菌中的常驻质粒。此外,由于其广泛的宿主范围,pKJK5::csg 成功阻断了一系列环境、猪和人类相关大肠杆菌分离株以及两种假单胞菌分离株中的 AMR 质粒摄取。这项研究坚定地确立了 pKJK5::csg 是一种有前途的广泛宿主范围 CRISPR-Cas9 递送工具,用于去除 AMR 质粒,它有可能应用于复杂的微生物群落,以从广泛的细菌物种中去除 AMR 基因。

频谱冠状病毒抗病毒药和可药物宿主目标
由许可药物组成的摘要图书馆代表了调节人类生理过程的大量分子曲目,为发现宿主靶向抗病人提供了独特的机会。我们筛选了重新利用,集中救援和加速的Medchem(倒置),以大约12,000个分子重新使用库,用于宽光谱冠状病毒抗病毒药,发现了134种化合物,抑制了αOronavirus并映射到58个分子靶标。主要的靶标包括5-羟基氨基胺受体,多巴胺受体和细胞周期蛋白依赖性激酶。Gene knock-out of the drugs' host targets including cathepsin B and L (CTSB/L; VBY-825), the aryl hydrocarbon receptor (AHR; Phortress), the farnesyl-diphosphate farnesyltransferase 1 (FDFT1; P-3622), and the kelch-like ECH-associated protein 1 (KEAP1; Omaveloxolone), significantly调节HCOV-229E感染,提供了证据表明这些化合物通过对各自的宿主靶标的作用抑制了病毒。对所有134个主要的化合物进行SARS-COV-2和验证的对识别的原始细胞的验证,AHR激活配体,P-3622靶向FDFT1和OmavelOxolone,以及Omaveloxolone,该a和Omaveloxolone激活NFE2样的BZIP转录因子2(nFe2L2),该nfe 2(nFe2L2)的kap kap and and and and and and and and and and and and them keap kap keap,kap and and and and and and and and the trib kap, alpha-和betacor onavirus。 这项研究提供了HCOV-229E重新利用候选者的概述,并揭示了被不同冠状病毒劫持的新型潜在可吸毒的病毒宿主依赖性因素。对识别的原始细胞的验证,AHR激活配体,P-3622靶向FDFT1和OmavelOxolone,以及Omaveloxolone,该a和Omaveloxolone激活NFE2样的BZIP转录因子2(nFe2L2),该nfe 2(nFe2L2)的kap kap and and and and and and and and and and and and them keap kap keap,kap and and and and and and and and the trib kap, alpha-和betacor onavirus。这项研究提供了HCOV-229E重新利用候选者的概述,并揭示了被不同冠状病毒劫持的新型潜在可吸毒的病毒宿主依赖性因素。
脊髓状宿主耗尽微生物DNA试剂盒
下列的PCR结果是使用小骨宿主耗尽微生物DNA试剂盒从唾液样品中提取DNA的结果,显示有效的宿主DNA耗竭和微生物DNA恢复。使用QPCR分析,据估计,对于这些样品的宿主DNA耗竭和细菌DNA恢复估计高于90%。图1:使用小骨宿主耗尽微生物DNA试剂盒从唾液样品中提取的DNA的PCR。a)使用人β-珠蛋白引物对宿主DNA检测。b)使用16S引物对细菌DNA检测。m:DNA标记;泳道1、3、5:提取的总DNA,没有执行宿主耗竭步骤;泳道2、4、6:宿主耗尽的(H. dep)DNA使用脊柱状宿主耗尽微生物DNA试剂盒提取;泳道7:PCR阴性对照。

分子基础和下游免疫后果的宿主相互作用
hal是一个多学科的开放访问档案,用于存款和传播科学研究文件,无论它们是否已发表。这些文件可能来自法国或国外的教学和研究机构,也可能来自公共或私人研究中心。

甲氨蝶呤用于移植与宿主疾病预防
在5至15分钟内50 mL氯化钠中0.9%(至少在干细胞输注后24小时)每天一次(非 - 刺激性)在每位甲氨蝶呤剂量+3,+6, +6,+6,+11 **确认为每种甲基疗法注册或顾问之前,用血液学注册官或顾问确认每种甲基甲酸剂或顾问,每种甲基甲基甲基甲酸甲酯剂量为50次甲基甲酸甲酸甲酸甲酸甲酯剂量,并在氯化钠在5至15分钟内每天一次0.9%(非 - 刺激性)频率:N/A(仅单剂量)循环数:1 *天1剂量的甲氨蝶呤为15mg/m 2或10mg/m 2。根据干细胞供体的来源,根据顾问的判断确定,使用抗胸腺细胞球蛋白在调节中使用抗心理细胞球蛋白和毒性风险(例如,粘膜炎和急性肾脏损伤)。参见参考文献,其中包括15mg/m 2和10mg/m 2的第1天甲氨蝶呤剂量。**第11天甲氨蝶呤剂量可以根据顾问的酌处权省略,具体取决于粘膜炎程度和其他毒性(例如急性肾脏损伤)的存在。在每个甲氨蝶呤剂量后24小时内可以考虑叶酸救援,因为它与毒性降低有关,并且不会增加GVHD或移植排斥的风险