XiaoMi-AI文件搜索系统
World File Search System抑制的


KMT2D 缺乏会促进易受核糖体生物合成抑制的髓系白血病
KMT2C 和 KMT2D 是人类癌症中最常见的突变表观遗传基因。虽然 KMT2C 被确定为急性髓系白血病 (AML) 中的肿瘤抑制因子,但 KMT2D 在这种疾病中的作用仍不清楚,尽管它的缺失会促进 B 细胞淋巴瘤和各种实体癌。据报道,KMT2D 在 AML 中下调或突变,并且通过 shRNA 敲低或 CRISPR/Cas9 编辑导致其缺陷会加速小鼠的白血病形成。造血干细胞和祖细胞以及 Kmt2d 缺失的 AML 细胞的核糖体生物合成显著增强,并且核仁持续增大,rRNA 和蛋白质合成率增加。从机制上讲,发现 KMT2D 缺陷会导致小鼠和人类 AML 细胞中 mTOR 通路的激活。 Kmt2d 直接调节 Ddit4 的表达,Ddit4 是 mTOR 通路的负调节因子。与核糖体生物合成异常一致,研究表明,RNA 聚合酶 I 抑制剂 CX-5461 可显著抑制体内 Kmt2d 缺失的 AML 生长,并延长白血病小鼠的生存期。这些研究证实 KMT2D 是 AML 中事实上的肿瘤抑制因子,并揭示了对核糖体生物合成抑制前所未有的脆弱性。

具有 1.7 dBm 输出功率和 45 dB 基波抑制的 E 波段 CMOS 频率四倍频器
摘要:本文介绍了一种采用40nm CMOS工艺的E波段四倍频器。该电路采用两个推推式倍频器和两个单级中和放大器。倍频器采用伪差分B类偏置共源共栅拓扑结构,提高了反向隔离度和转换增益。采用中和技术可同时提高放大器的稳定性和功率增益。堆叠变压器用于单端到差分转换以及输出带通滤波。输出带通滤波器可提高四次谐波的输出功率,同时抑制不需要的谐波,特别是二次谐波。核心芯片尺寸为0.23mm2,功耗为34mW。测得的四次谐波在76GHz时实现了1.7dBm的最大输出功率,峰值转换增益为3.4dB。对于 74 至 82 GHz 的频谱,基波和二次谐波抑制分别超过 45 dB 和 20 dB。

独立于 RAS 的生物标志物组可可靠地预测结直肠癌对 MEK 抑制的反应
1 CELLphenomics GmbH,罗伯特-罗斯勒-Str。 10, 13125 柏林, 德国; ulrike.pfohl@cellphenomics.com(上); juergen.loskutov@cellphenomics.com (JL); christoph.reinhard@cellphenomics.com (CR); lena.wedeken@cellphenomics.com (LW) 2 法兰克福歌德大学分子生物科学研究所,Max-von-Laue-Str。 13, 60438 法兰克福, 德国 3 基因组工程与疾病模型,Max Delbrück 分子医学中心,Robert-Rössle-Str. 13, 60438 法兰克福, 德国10, 13125 柏林, 德国; sanum.bashir@biontech.de (SB); ralf.kuehn@mdc-berlin.de (RK) 4 NMI 自然科学与医学科学研究所,图宾根大学,Markwiesenstraße 55, 72770 Reutlingen,德国;patrick.herter@boehringer-ingelheim.com (PH);markus.templin@nmi.de (MT) 5 ASC Oncology GmbH,Robert-Rössle-Str. 10, 13125 Berlin,德国 6 病理学研究所,柏林夏利特大学医学院,Virchowweg 15, Charité pl. 1, 10117 Berlin,德国;soulafa.mamlouk@charite.de 7 生物技术研究所,赫尔辛基大学,Viikinkaari 5, Biocenter 2, 00790 Helsinki,芬兰; sergei.belanov@helsinki.fi 8 罗斯托克大学医学中心普通外科、分子肿瘤学和免疫治疗诊所,Schillingallee 35, 18057 罗斯托克,德国;michael.linnebacher@med.uni-rostock.de 9 罗斯托克大学医学中心普通外科诊所,Schillingallee 35, 18057 罗斯托克,德国;florian.buertin@med.uni-rostock.de 10 巴塞尔大学医院,Petersgraben 4, 4031 巴塞尔,瑞士;marcus.vetter@ksbl.ch 11 巴塞尔兰州立医院,Rheinstr. 26, 4410 Liestal, Switzerland 12 Institute of Pathology, University Hospital Göttingen, Robert-Koch-Straße 40, 37075 Göttingen, German * 通讯地址:christian.regenbrecht@cellphenomics.com

靶向 BCL2 可克服小细胞肺癌的耐药性并增强 AZD2811 对 Aurora 激酶 B 抑制的反应
利益冲突声明 LAB 获得阿斯利康和安进的研究支持,并担任默克夏普和多姆公司、Arrowhead Pharmaceuticals、Chugai Pharma、阿斯利康、Genetech Inc.、Abbvie、BeiGene、Jazz Pharmaceuticals 的咨询/顾问。CMG 是阿斯利康、百时美施贵宝、Jazz Pharmaceuticals 和 MonteRosa 的顾问委员会成员,也是阿斯利康、百时美施贵宝和 Jazz Pharmaceuticals 的演讲局成员。CMG 还获得了阿斯利康的研究支持。CMD 获得了安进和阿斯利康的差旅补助,以及罗氏、默沙东和默克的个人演讲费。在生成本稿件的数据时,UP、CA、JS、JEPJT、GF 和 JU 是阿斯利康的员工和股东。所有其他作者声明没有竞争利益。

Mammaprint®与基因表达途径的关联预测对细胞周期蛋白依赖性激酶4/6(CDK)抑制的抗性
•在所有HR+HER2-肿瘤中,14.4%(n = 834)被归类为Ultralow,38.1%为(n = 2201)低,37.6%(n = 2175)为高1,而9.8%(n = 568)为高2风险(表1)。•观察到线性相关性,随着哺乳动物的风险增加和RB功能的增加(p <0.001)(图2)。RB功能丧失的平均基因表达与高2个肿瘤表现出最高的相关性。所有哺乳动物/蓝图亚型比较都是显着的(所有比较的2路ANOVA,p <0.0001)(图3)。•与其他哺乳动物组相比,乳腺斑点高2的肿瘤比例最高,对CDK4I的耐药性(43.0%)(p <0.001)(图4)。•高2显示与与CDK4无关的高增殖相关的基因最强的相关性,与所有哺乳动物基团相比,相关性明显更高(所有乳突比较的2条ANOVA,P <0.0001)(图5A)。然而,在蓝图高2个腔与高2基底亚组比较之间对CDK4I的抗性之间的相关性中没有显着差异(2路ANOVA,P = 0.185)(图5B)。

荟萃分析供体衍生的无细胞DNA作为肾脏同种移植抑制的诊断标记:系统评价和荟萃分析
供体衍生的无细胞DNA(DD-CFDNA)已成为检测移植排斥反应的有前途的生物标志物。这项研究旨在评估将其应用于肾脏移植排斥的诊断准确性和临床价值。从PubMed,Embase,Cochrane Library和Web of Science数据库中审查了有关肾脏移植拒绝中DD-CFDNA诊断的相关文献。数据和研究特征由两名研究人员独立提取。分别分析了任何排斥(AR)和抗体介导的排斥反应(ABMR)的诊断精度数据。潜在的异质性。漏斗图用于阐明出版偏见的存在或不存在。九本出版物提供了有关诊断AR患者的DD-CFDNA准确性的数据。具有95%置信区间(CIS)的接收器操作特征(AUROC)曲线下的汇总敏感性,特异性和面积为0.59(95%CI,0.48 - 0.69),0.83(95%CI,0.76 - 0.88)和0.80(95%CI,0.80(95%CI,0.76 - 0.76 - 0.83)。此外,12项研究集中在ABMR的DD-CFDNA的诊断准确性上,显示了95%CI为0.81(95%CI,0.72 - 0.88),0.80(95%CI,0.73 - 0.73 - 0.86)和0.87(95%)和0.87(95%)(95%CI,0.87(95%),95%CI(95%CI,0.72 - 0.88)和0.87(95%CI,0.84%),0.84(95%CI),表明汇总的灵敏度,特异性和AUROC曲线。研究类型,年龄组和样本量导致异质性。总而言之,我们的发现表明,虽然血浆DD-CFDNA诊断AR患者的准确性受到明显的异质性的限制,但它是诊断ABMR的有价值的生物标志物。

在乳腺腔腔上皮细胞中具有致癌潜力表观遗传抑制的表观遗传抑制
哺乳动物基因组中DNA甲基化的主要功能是抑制转座元素(TES)。在癌细胞中通常观察到的广泛的甲基化损失导致TE的表观遗传抑制丧失。衰老过程的特征是甲基甲基的变化。然而,这些表观基因组改变对沉默的影响及其功能后果尚不清楚。为了评估衰老中TES的表观遗传调节,我们在人类乳腺腔上皮细胞(LEPS)中介绍了DNA甲基化(LEPS),这是一种与年龄较大的乳腺癌有关的关键细胞谱系 - 来自年龄较大的乳腺癌。我们在这里报告说,几个TE亚家族在正常LEP中充当调节元素,并且这些子集的一部分显示出随着年龄的增长而显示一致的甲基化变化。在这些TES处的甲基化变化发生在谱系特异性转录因子结合位点,与谱系特异性的丧失一致。主要显示甲基化损失,而CpG岛(CGI)是Polycomb抑制性复合物2(PRC2)的靶标,显示衰老细胞中甲基化的增加。在衰老的LEP中,许多具有甲基化损失的TE都有乳腺癌样品中调节活性的证据。我们还表明,TES的甲基化变化会影响与腔乳腺癌相关的基因的调节。这些结果表明,衰老会导致TES的DNA甲基化变化,从而弥补了维持谱系特异性,并可能增加对乳腺癌的敏感性。
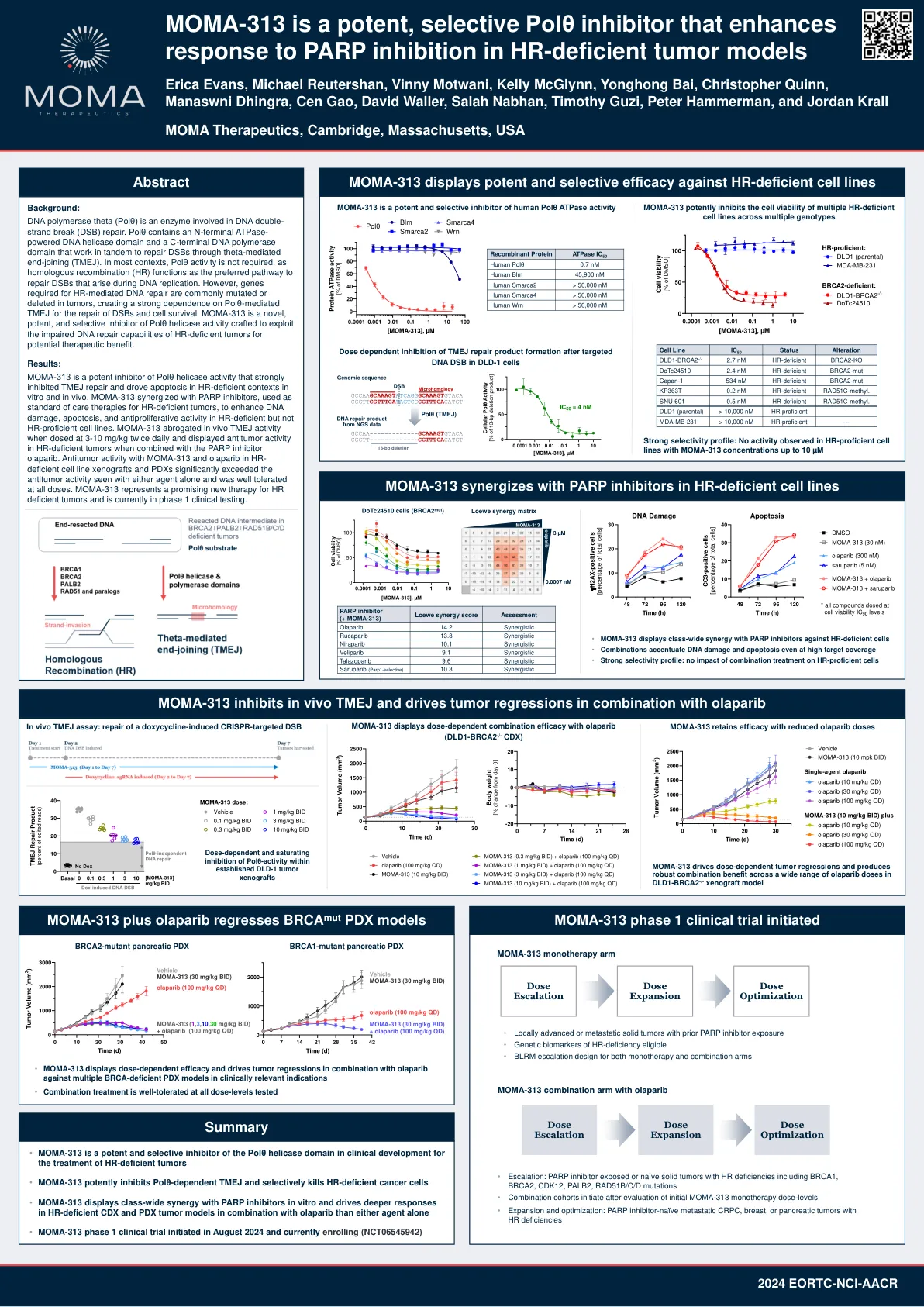
MOMA-313 是一种强效、选择性 Polθ 抑制剂,可增强 HR 缺陷肿瘤模型对 PARP 抑制的反应
DNA 聚合酶 theta (Polθ) 是一种参与 DNA 双链断裂 (DSB) 修复的酶。Polθ 包含一个 N 端 ATPase 驱动的 DNA 解旋酶结构域和一个 C 端 DNA 聚合酶结构域,它们协同作用,通过 theta 介导的末端连接 (TMEJ) 修复 DSB。在大多数情况下,Polθ 活性不是必需的,因为同源重组 (HR) 是修复 DNA 复制过程中出现的 DSB 的首选途径。然而,HR 介导的 DNA 修复所需的基因通常在肿瘤中发生突变或缺失,导致 DSB 修复和细胞存活严重依赖 Polθ 介导的 TMEJ。MOMA-313 是一种新型、有效且选择性的 Polθ 解旋酶活性抑制剂,旨在利用 HR 缺陷型肿瘤受损的 DNA 修复能力来获得潜在的治疗益处。

立场文件 释放南非电网容量,实现快速可再生能源部署 可再生能源拥堵抑制的重要作用
1 资料来源:IPP 办公室 www.ipp-renewables.co.za 2 资料来源:IEA 2023 年电力市场报告:https://www.iea.org/reports/electricity-market-report-2023/executive-summary 3 50Hertz Transmission 是德国四家 TSO 之一,为该国东北部的 1800 万客户提供服务,年总负荷约为 100 TWh。其中三分之二的能源由 50Hertz 电网区域内的 RES 提供。 4 资料来源:50Hertz 2022 年事实和数据报告:https://www.50hertz.com/xspProxy/api/staticfiles/50hertz-client/doku-mente/medien/publikationen/50hertz_facts_and_figures2022.pdf